.
Condición fitosanitaria: Presente
Grupo de cultivos: Frutales de pepita
Especie hospedante: Peral (Pyrus communis)
Rango de hospedantes: específico / estrecho. Infecta Pyrus communis y algunas ornamentales emparentadas (Eriobotrya japonica, etc.). (*)
Etiología: Hongo. Necrotrófico (considerado hemibiotrófico en una fase inicial)
Agente causal: Venturia pyrina (teleomorfo), Fusicladium pyrorum (Lib.) Fuckel [as ‘pirinum’], (1870) (anamorfo)
Taxonomía: Eukaryota > Fungi > Dikarya > Ascomycota > Pezizomycotina > Dothideomycetes > Dothideomycetes incertae sedis > Venturiales > Venturiaceae > Venturia
.
-

- Autor: Dra. Cecilia Lutz
.
(*) Las infecciones de Venturia spp. se limitan a seis familias de plantas hospedantes: Acaraceae, Betulaceae, Cornaceae, Oleaceae, Rosaceae y Salicaceae (Sivanesan, 1977). Las especies de Venturia spp. también parecen ser muy específicas del hospedante en el sentido de que cada especie suele estar confinada a un género de planta o al menos a géneros hospedantes afines dentro de una familia (Schubert et al., 2003). Sin embargo, existe cierta controversia con respecto al rango de hospedantes de Venturia spp. en árboles frutales. En la monografía de Venturia publicada en 1977, Sivanesan enumeró 14 especies de Rosaceae como hospedantes de V. inaequalis, y estos hospedantes pertenecían a los géneros Cotoneaster, Malus, Pyracantha, Pyrus y Sorbus. Posteriormente, Schubert et al. (2003) aumentaron el número de hospedantes de V. inaequalis para incluir 12 géneros, agregando Amelanchier, Aronia, Docynia, Eriobotrya, Heteromeles, Kageneckia y Prunus. Sin embargo, estos últimos autores no citaron estudios específicos sobre la capacidad de V. inaequalis para infectar a estos hospedantes, y algunas de estas interacciones hospedante-patógeno han sido refutadas (Menon, 1956; Raabe y Gardner, 1972; Ishii y Yanase, 2000; Stehmann et al., 2001; Le Cam et al., 2002; Chevalier et al., 2004; Sánchez-Torres et al., 2007, 2009; Abe et al., 2008). Otros estudios, no han logrado obtener infección de pera por V. inaequalis o infección de manzana por V. pyrina (Menon, 1956; Stehmann et al., 2001; Chevalier et al., 2004). Menon (1956) inoculó plantas de manzano y peral con tres tipos de inóculo diferentes tanto de V. inaequalis como de V. pyrina (micelio, suspensiones de conidios y suspensiones de ascosporas) y observó síntomas claros de sarna en manzanos solo con V. inaequalis y en perales solo con V. pyrina, y en ambos casos sólo con conidios o ascosporas. Stehmann et al. (2001) encontraron pseudotecios tanto de V. inaequalis como de V. pyrina en hojas muertas de manzano, pero las inoculaciones cruzadas con las suspensiones de conidios de estos aislamientos en hojas desprendidas de manzano y peral dieron como resultado una infección solo cuando se inoculó V. inaequalis en el manzano y cuando se inoculó el peral con V. pyrina. Aunque los conidios de V. inaequalis y V. pyrina germinaron en ambos hospedantes, y aunque los tubos germinativos produjeron apresorios e hifas ectotróficas en ambos hospedantes, solo se produjo una densa red subcuticular de estroma en el hospedante compatible (Stehmann et al., 2001); Chevalier et al., 2004). Las plantas de níspero (E. japonica) inoculadas con V. pyrina mostraron síntomas en las hojas, pero los síntomas difirieron de los causados por el hongo de la sarna del níspero F. eriobotryae (Sánchez-Torres et al., 2007). Raabe y Gardner (1972) infectaron con éxito plantas de níspero con F. pyracanthae. En forma global, los resultados de las inoculaciones cruzadas de Venturia spp. en diferentes hospedantes rosáceos muestran que estos patógenos son específicos del hospedante, y la información contenida en las monografías de Schubert et al. (2003) y Sivanesan (1977) no se pueden reproducir. La especificidad por el hospedante requiere más estudio para F. pyracanthae y F. eriobotryae.
.
.
.
Antecedentes
La sarna del peral es una enfermedad grave en todas las regiones productoras de pera del mundo (Spotts y Cervantes, 1994; Dobra et al., 2007). La enfermedad destruye la calidad comercial de los frutos y el rendimiento de las plantas. Se han llegado a registrar pérdidas que alcanzaron entre 40 y 80% (Liu et al., 2009). En el Alto Valle, su ocurrencia en el Alto Valle está muy ligada a las condiciones ambientales, con presencia esporádica, asociada a primaveras lluviosas (Dobra et al., 2007). La incidencia de la enfermedad depende de las variedades de peral plantadas y de las condiciones climáticas estacionales y microclimáticas de cada monte. En los últimos años se han observado elevados niveles de infección, comprometiendo la productividad en pera de la variedad William´s, y en algunos casos hasta pérdidas totales de la producción (Sosa y Lutz, 2020).
.
Síntomas
Los síntomas son muy semejantes en manzano y peral y pueden aparecer en todos los órganos aéreos de la planta, excepto en las partes lignificadas del árbol. Desde el estado fenológico de yema hinchada, las hojas, las flores (excepto pétalos) y los frutos (hasta que alcanzan el tamaño de una nuez), son susceptibles a la infección. Cuando ocurren condiciones ambientales conducentes a la infección, se observan los síntomas entre los 7 – 10 días (incubación) en el envés de hojas, sépalos, pedúnculo y receptáculo de fruto. En fruta, las lesiones son deprimidas, negras, de bordes irregulares y escamosos. Las lesiones ocasionadas por infección ascospórica son de tamaño grande, mientras que las originadas por infección conídica son pequeñas (Sosa y Lutz, 2020).
.
.
-

- Lesiones de la sarna del peral en frutos, que provocan deformaciones. Autor: Bruce Watt

-

- Manchas oscuras (lesiones) de la sarna del peral provocadas por Venturia pyrina, mientras que las manchas pequeñas más claras son solo lenticelas. Autor: Neil Bell.


-

- Infecciones primarias y secundarias en frutos de pera Packham’s Triumph. Autor: Dra. Cristina Sosa

-

- Infecciones secundarias en frutos de pera Packham’s Triumph. Autor: Dra. Cristina Sosa

-

- Infecciones primarias y secundarias a cosecha en pera Packham’s Triumph. Autor: Dra. Cristina Sosa

-

- Síntomas de sarna del peral en hoja y fruto en crecimiento. Autor: Dra. Cristina Sosa

-

- Síntomas de sarna del peral en fruto en conservación. Autor: Dra. Cristina Sosa

-

- Síntomas de sarna del peral, infecciones primarias a campo. Autor: Dra. Cristina Sosa

-

- Síntomas de sarna del peral en fruto, infecciones primarias y secundarias. Autor: Dra. Cristina Sosa

.
.
Ciclo de la enfermedad y Epidemiología
Venturia pyrina sobrevive como pseudotecios en las hojas infectadas que caen al suelo del monte frutal y como conidios en las lesiones de las ramitas. La etapa de supervivencia más importante del microorganismo se produce como pseudotecio inmaduro en los tejidos del mesófilo de las hojas sobre el suelo, por lo tanto las mismas constituyen la fuente de inóculo primario. En la primavera, el hongo comienza su ciclo de vida arrojando sus ascosporas a través de las aberturas (ostíolo). Las ascosporas son el inóculo primario que dan comienzo a la enfermedad o ciclo primario. Las corrientes de aire las transportan hasta las hojas o frutos del manzano, más susceptibles en su juventud. Cuando las esporas disponen de humedad foliar se produce la germinación, los tubos germinativos que emiten las ascosporas atraviesan la cutícula sin que en apariencia exista un fenómeno de solubilización de ésta y el micelio comienza a crecer formando un estroma subcuticular. Pocos días después de la infección numerosos conidióforos cortos irrumpen a través de la cutícula y cada uno produce en su extremo un conidio. Los conidios se dispersan por las lluvias y llegan a otras hojas o frutos jóvenes. El hongo se propaga asexualmente produciendo varias generaciones conidiales, por lo tanto los conidios o esporas asexuales son el inóculo secundario siendo las lesiones del año la fuente de inóculo secundario, en esta etapa (ciclo secundario) los conidios renuevan el ciclo de la enfermedad durante la primavera y el verano produciendo las infecciones secundarias tantas veces como se den las condiciones climáticas predisponentes.
Ya avanzada la estación de crecimiento, cuando las células de las hojas comienzan a morir, el micelio penetra profundamente dentro de los tejidos. El desarrollo de los pseudotecios, ascos y ascosporas continúa siendo regulado por factores tales como la temperatura y la humedad.
.
-
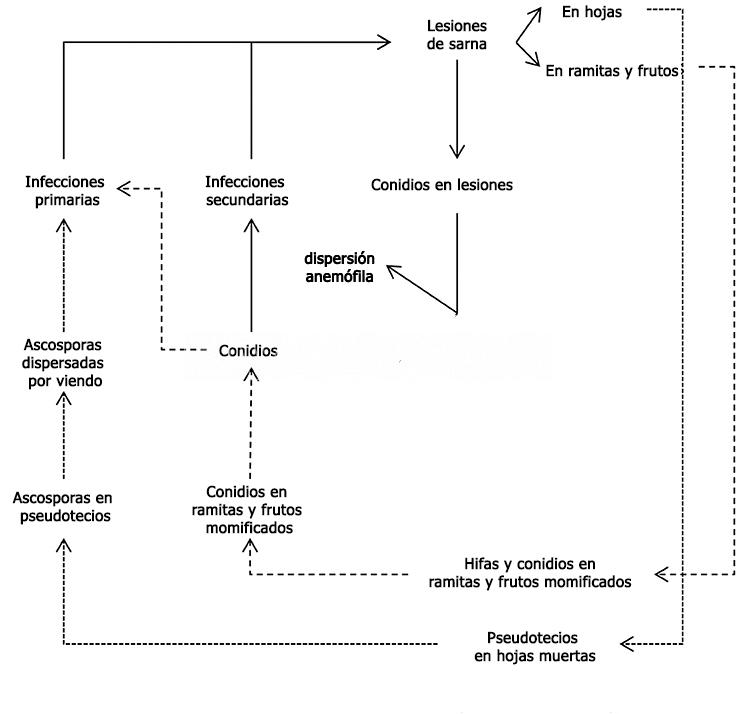
- Ciclo de vida de Venturia spp. Las líneas punteadas (…) indican la fase sexual del ciclo de vida; las líneas discontinuas (—–) indican la fase asexual del ciclo de vida. Autor: González-Domínguez et al., 2017
.
.
Manejo Integrado
El control de la enfermedad se basa en un modelo epidemiológico diseñado para la sarna del manzano (causada por V. inaequalis).
.
.
.
Bibliografía
Chevalier M, Bernard C, Tellier M, et al. (2004). Host and non-host interaction of Venturia inaequalis and Venturia pirina on Pyrus communis and Malus x domestica. Acta Hortic. 663: 205–208. doi: 10.17660/ActaHortic.2004.663.30
Dobra A, Rossini M, Barnes N, Sosa C (2007) Manejo integrado de enfermedades de los frutales de pepita. En: Árboles frutales: Ecofisiología, Cultivo y Aprovechamiento. Sozzi (ed.) 1ªed. FAUBA, 587-615.
Eikemo H, Gadoury D, Spotts R, et al. (2011) Evaluation of six models to estimate ascospore maturation in Venturia pyrina. Plant Disease 95: 279-284. doi: 10.1094/PDIS-02-10-0125
González-Domínguez E, Armengol J, Rossi V (2017) Biology and Epidemiology of Venturia Species Affecting Fruit Crops: A Review. Frontiers in Plant Science 8: 1496. doi: 10.3389/fpls.2017.01496
Ishii H, Yanase H (2000) Venturia nashicola, the scab fungus of Japanese and Chinese pears: a species distinct from Venturia pirina. Mycol. Res. 104: 755–759. doi: 10.1017/S0953756299001720
Johnson S, Jones D, Thrimawithana AH, et al. (2019) Whole Genome Sequence Resource of the Asian Pear Scab Pathogen Venturia nashicola. Mol Plant Microbe Interact. 32(11): 1463-1467. doi: 10.1094/MPMI-03-19-0067-A
Le Cam B, Sargent D, Gouzy J, et al. (2019) Population Genome Sequencing of the Scab Fungal Species Venturia inaequalis, Venturia pirina, Venturia aucupariae and Venturia asperata. G3 (Bethesda) 9(8): 2405-2414. doi: 10.1534/g3.119.400047
Menon R (1956) Studies on Venturiaceae on rosaceous plants. Phytopathol. Z. 27: 117–146.
Raabe R, Gardner MW (1972) Scab of pyracantha, loquat, toyon and kageneckia. Phytopathology 62: 914–916. doi: 10.1094/Phyto-62-914
, , , et al. (2022) A new highly effective hybrid fungicide containing difenoconazole and tea tree oil for managing scab of apple, pecan and almond trees and as a tool in resistance management. Plant Pathology 71: 1774– 1783. doi: 10.1111/ppa.13610
Sánchez-Torres P, Hinarejos R, Tuset JJ (2007) Fusicladium eriobotryae: hongo causante del moteado del níspero en el Mediterráneo español. Bol. San. Veg. Plagas 33: 89–98.
Sánchez-Torres P, Hinarejos R, Tuset JJ (2007) Identification and characterization of Fusicladium eriobotryae: fungal pathogen causing mediterranean loquat scab. Acta Hortic. 33, 343–347. doi: 10.17660/ActaHortic.2007.750.54
Sánchez-Torres P, Hinarejos R, Tuset JJ (2009) Characterization and pathogenicity of Fusicladium eriobotryae, the fungal pathogen responsible for loquat scab. Plant Disease 93: 1151–1157. doi: 10.1094/PDIS-93-11-1151
Schubert KS, Ritschel AR, Braun UB (2003) A monograph of Fusicladium s. lat. (Hyphomycetes). Schlechtendalia 9: 1–132.
Sivanesan A (1977) The Taxonomy and Pathology of Venturia Species. Bibliotheca Mycologica. Vaduz: Lubrecht and Cramer Ltd.
Sosa MC, Lutz MC (2020) Enfermedades del peral (Pyrus communis) en montes comerciales de la norpatagonia argentina. Boletín de la Asociación Argentina de Fitopatologos N°6. Link
Spotts A, Cervantes A, Niederholzer A (1997) Effect of dolomitic lime on production of asci and pseudothecia of Venturia inaequalis and V. pirina. Plant Disease 81: 96-98. doi: 10.1094/PDIS.1997.81.1.96
Stehmann C, Pennycook S, Plummer KM (2001) Molecular identification of a sexual interloper: the pear pathogen, Venturia pirina, has sex on apple. Phytopathology 91: 633–641. doi: 10.1094/PHYTO.2001.91.7.633
.