.
Condición fitosanitaria: Plaga no cuarentenaria reglamentada
Grupo de cultivos: Frutales (Cítricos)
Rango de hospedantes: específico / estrecho (Cítricos)
Especie hospedante:
…* Limón (Citrus limon (L.) Burm.f.),
…* Mandarina (Citrus reticulata Blanco),
…* Naranjo (Citrus sinensis (L.) Osbeck),
…* Pomelo (Citrus × paradisi Macfad.),
…* tangor (Citrus nobilis Lour.)
.
Grado de susceptibilidad:
…* Muy susceptible: pomelos, lima Key
…* Susceptible: naranjas tempranas, mandarinas híbridas, mandarina clementina, limón
…* Resistente: naranja Valencia, algunas mandarinas
…* Muy resistente: mandarinas Okitsu, lima Tahití
…* No se infectan a campo: kumquat (falso quinoto)
.
Epidemiología: subaguda, policíclica (progreso intraanual), poliética (progreso interanual).
Etiología: Bacteria. Gram negativa. Considerada hemibiótrofa
Agente causal: Xanthomonas citri subsp. citri = Xanthomonas citri pv. citri (*) (ex Hasse 1915) Gabriel et al. 1989 emend. Constantin et al. 2016
y Xanthomonas fuscans subsp aurantifolii (Xfa) en pomelo (ver Ference et al., 2018)
Taxonomía: Bacteria > Proteobacteria > Gammaproteobacteria > Xanthomonadales > Xanthomonadaceae > Xanthomonas > Xanthomonas citri group
.
(*) Dado que la cancrosis de los cítricos se describió por primera vez a principios de 1900, se identificaron cuatro biotipos principales y tres especies de Xantomonas patógenas asociadas a los cítricos en función de los síntomas característicos y el rango de hospedantes. El biotipo A, causado por X. citri subsp. citri, desarrolla lesiones elevadas típicas de pseudocancro y es patógeno en la mayoría de las especies y cultivares de cítricos comerciales. Los biotipos B y C son causados por X. citri pv. aurantifolii que provoca lesiones de pseudocancro. Además del biotipo típico A, el biotipo más agresivo y más extendido, se han descrito otros subtipos, como el biotipo A *, biotipo Aw, biotipo A-Manatí, biotipo A-Miami, biotipo A-menos pthA y biotipo-A etrog. Las cepas de biotipo B se encuentran solo en Argentina y son más patógenas en limón y menos agresivas en naranja dulce, mandarinas y pomelo. Las cepas de biotipo C se encuentran solo en Brasil y son patógenas solo en la lima Key. X. alfalfae subsp. citrumelonis, anteriormente conocido como biotipo E, causa manchas bacterianas en los cítricos y es más agresivo en los cítricos trifoliados y sus híbridos. Las lesiones causadas por la mancha bacteriana de los cítricos son planas o hundidas y no están elevadas. También se han identificado varios subtipos de X. citri subsp. citri (Patané et al., 2019). Además de las diferencias en patogenicidad, las xanthomonads también pueden variar en su sensibilidad al cobre (Behlau et al., 2020).
.
TIPOS DE CANCROSIS (según biotipo)
-Biotipo A o Cancrosis Asiática (Xanthomonas citri pv. citri): forma más virulenta que afecta a la mayoría de los cítricos; se encuentra en muchos países del mundo, entre ellos Argentina.
-Biotipo B o Sudamericana (Xanthomonas axonopodis pv. aurantifolli): aparición esporádica sólo en limoneros en Argentina (1927-29), denominada “falsa cancrosis” o “cancrosis sudamericana”.
-Biotipo C o Cancrosis de la Lima Key (Xanthomonas axonopodis pv. aurantifolli): apareció en Florida (EE.UU.; 1984) sobre citrumelo. No es una enfermedad seria.
-Biotipo D (México)
-Biotipo E, Cancrosis de vivero o mancha bacteriana de los cítricos (Xanthomonas axonopodis pv. citrumelo): Se halló sólo en Brasil (1960) afectando lima Key (Citrus aurantifolia) siendo de ocurrencia esporádica. No causa cancros, sino manchas.
.
Distintas especies de Xanthomonas de dos grupos filogenéticos son los agentes causantes del cancro de los cítricos. El grupo asiático, representado por Xanthomonas citri subsp. citri (X. citri tipo “A”) se distribuye ampliamente, tiene un amplio rango de hospedantes y es responsable de los síntomas más graves de cancro. El grupo sudamericano tiene un rango de hospedantes más restringido y comprende dos cepas de Xanthomonas fuscans subsp. aurantifolii (tipos: “B” y “C”) . El patotipo «C» provoca una reacción de hipersensibilidad en la mayoría de las especies de cítricos, causando enfermedad solo en Citrus aurantifolii (lima mexicana), mientras que el patotipo «B», aunque es capaz de causar cancros leves en naranjas, muestra síntomas más graves en limón (Citrus limon) y lima mexicana (Citrus aurantifolii) (Martins et al., 2020).
.
-
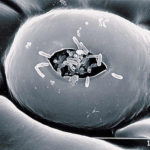
- SEM (scanning electron microscope) de estomas en hoja de pomelo con bacterias Xanthomonas campestri pv. citri (Xcc) entrando en la cámara estomática. Autor: J. Cubero.

-
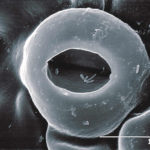
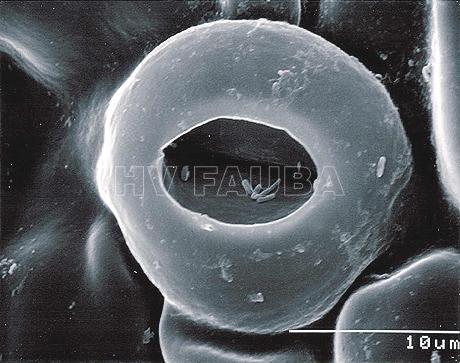
- SEM (scanning electron microscope) de estomas en hoja de pomelo con bacterias Xanthomonas campestri pv. citri (Xcc) en la cámara estomática. Autor: J. Cubero.
-
- Autor: Florida Division of Plant Industry , Florida Department of Agriculture and Consumer Services, Bugwood.org

.
.
.
Antecedentes
La cancrosis de los cítricos (cancro de los cítricos) existe desde tiempos remotos en Asia y desde allí se diseminó al resto del mundo. Afecta a casi todos los cítricos (Citrus spp.) y especies relacionadas con este género dentro de la familia Rutaceae. Pomelo y naranjo trifolio son altamente susceptibles; limonero, los naranjos dulce y agrio, moderadamente susceptibles y los mandarinos, moderadamente resistentes.
La cancrosis se reportó en Argentina desde 1928 como la forma de tipo B. La baja agresividad y el rango restringido de hospedantes limitaron las cepas de tipo B durante 40 años a un área pequeña. Estas cepas desaparecieron en 1978-90 después de la introducción de las cepas más agresivas de tipo A en 1974. La sustitución de las cepas de tipo B con las cepas de tipo A pudo haber tenido lugar debido a la producción de sustancia de tipo bacteriocina (toxina proteica sintetizada por una bacteria con el fin de inhibir el crecimiento de bacterias similares o de cepas cercanas) por las cepas A contra las cepas B. Las cepas de cancro tipo B infectaron casi exclusivamente a limón y se obtuvo un control adecuado mediante aplicaciones de bactericidas que contienen cobre. El tipo A de cancro de los cítricos, causado por Xanthomonas citri tipo A (Xc-A; Xnthomonas axonopodis pv citri) se volvió endémico en la parte noreste de Argentina después de que los esfuerzos de erradicación fracasaron. El tipo A ingresó al NE en 1974, se propagó rápidamente de 1977 a 1980 y luego a una tasa constante hasta 1990, cuando se volvió endémico en la región. La presencia de cancro en el noroeste de Argentina fue evidente en 2002.
-
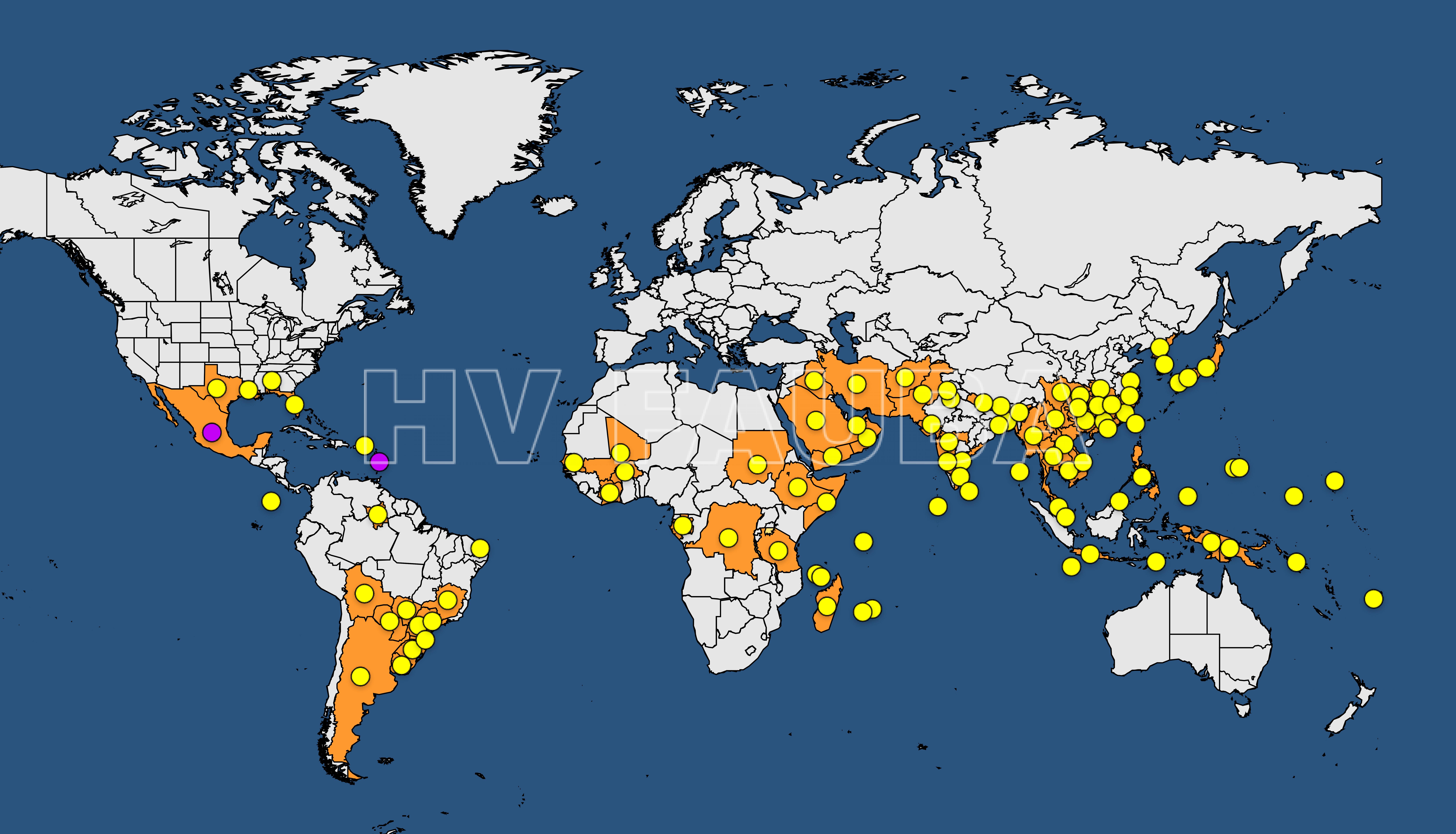
- Distribución mundial de la cancrosis de los cítricos. Fuente: EPPO Global Database: https://gd.eppo.int/taxon/XANTCI/distribution
.
Sintomatología
Los síntomas se manifiestan en casi toda la parte aérea del frutal: hojas, espinas, ramas jóvenes y adultas y frutos.
.
-

- Cancrosis del limonero. Síntomas en hoja (pseudocancro). Autor: Dra. Verónica Felipe
-
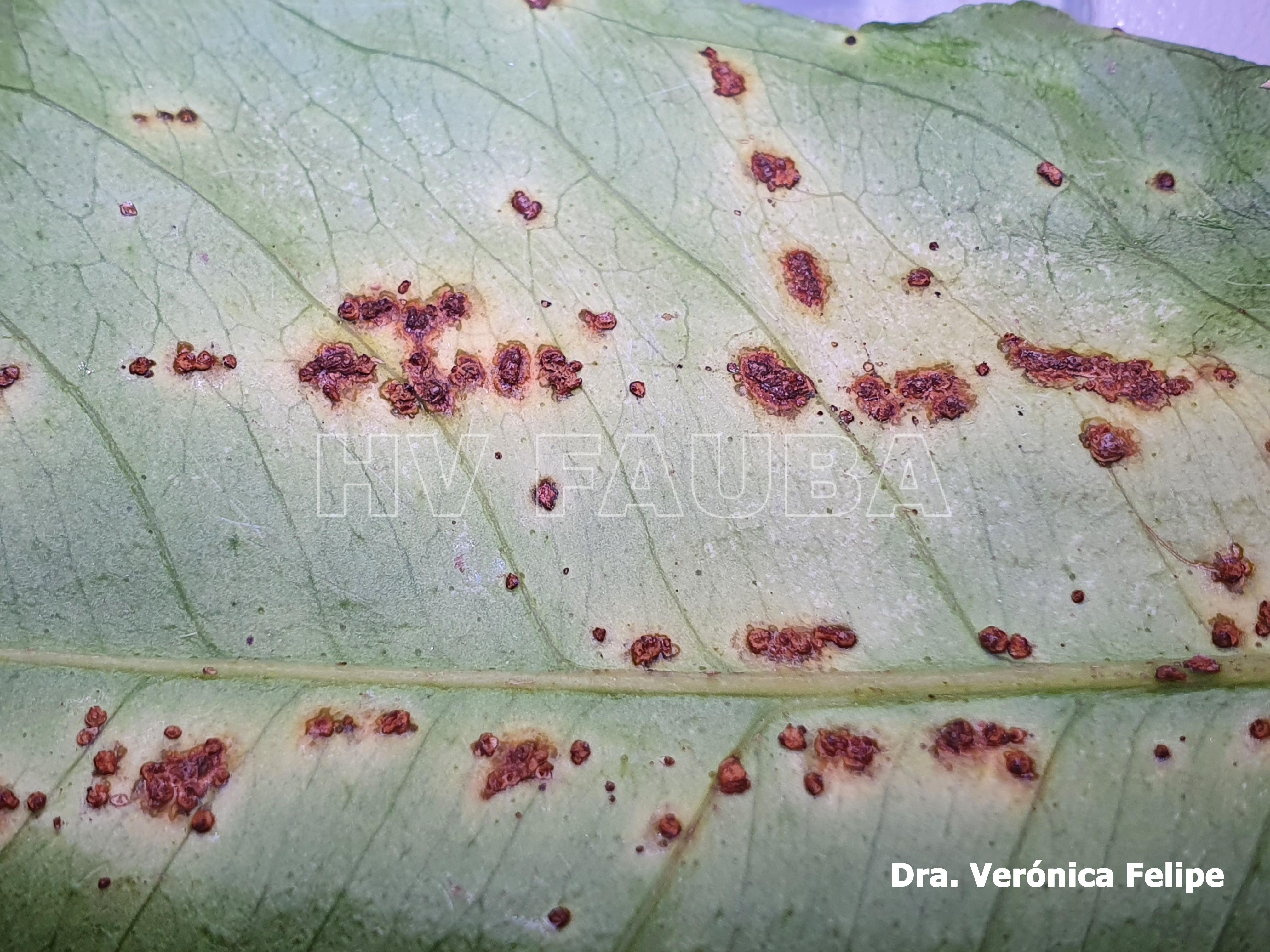
- Cancrosis del limonero. Síntomas en hoja (pseudocancro). Autor: Dra. Verónica Felipe
-
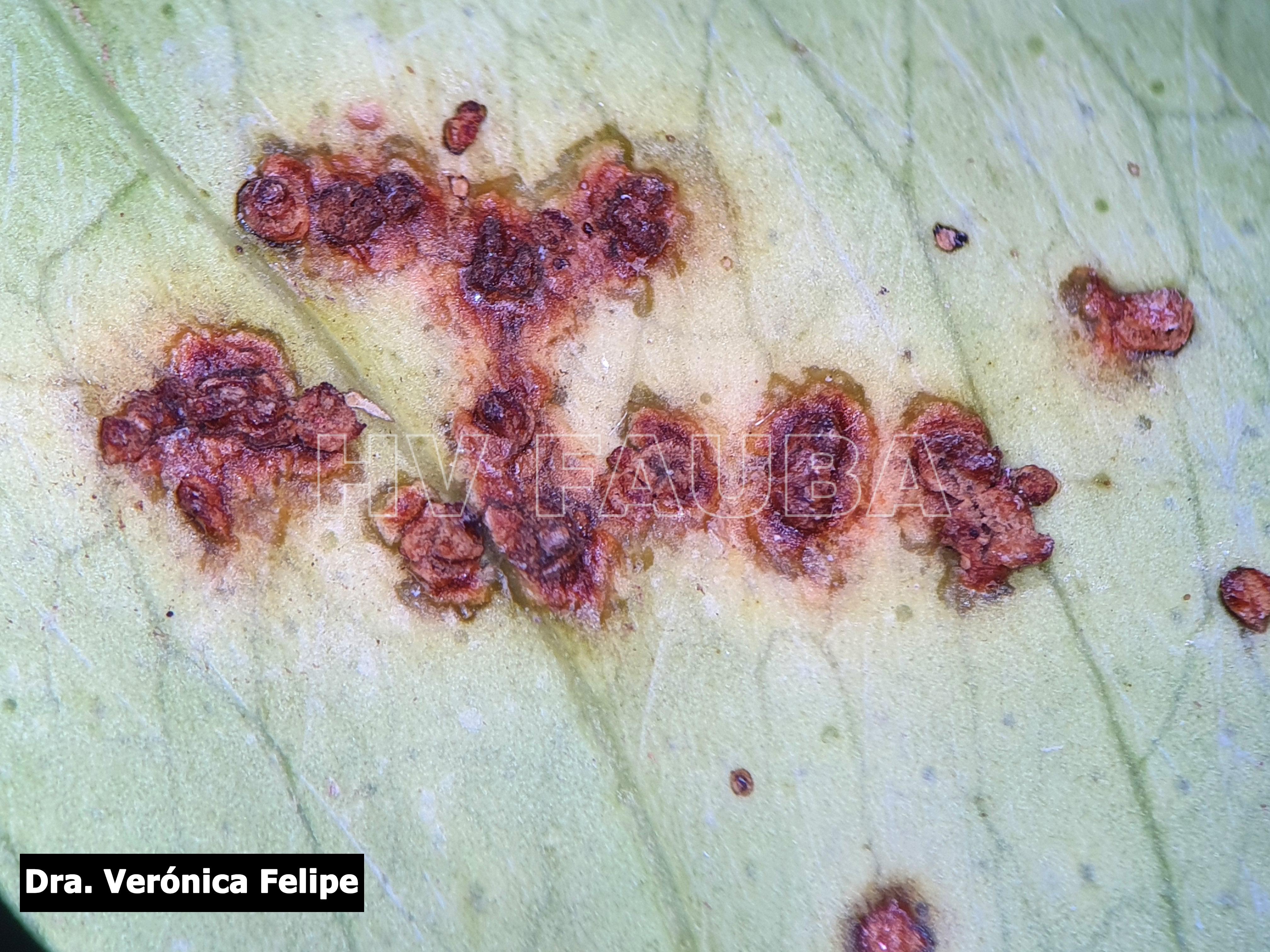
- Cancrosis del limonero. Síntomas en hoja (pseudocancro). Autor: Dra. Verónica Felipe
-

- Cancrosis del limonero. Síntomas en hoja (pseudocancro). Autor: Dra. Verónica Felipe
-

- Cancrosis del limonero. Síntomas en hoja (pseudocancro). Autor: Dra. Verónica Felipe
-

- Cancrosis del limonero. Síntomas en hoja (pseudocancro). Autor: Dra. Verónica Felipe
-

- Cancrosis del limonero. Síntomas en hoja (pseudocancro). Autor: Dra. Verónica Felipe
-

- Cancrosis del limonero. Síntomas en tallo (cancro). Autor: Dra. Verónica Felipe
-

- Cancrosis del limonero. Síntomas en tallo (cancro). Autor: Dra. Verónica Felipe
-

- Cancrosis del limonero. Síntomas en tallo (cancro). Autor: Dra. Verónica Felipe
-

- Autor: Claudio Baron
-

- Autor: Claudio Baron
-

- Autor: Claudio Baron
-

- Autor: Claudio Baron
-

- Autor: Claudio Baron
-

- Autor: USDA
-

- Autor: Jonas Janner Hamann, Universidade Federal de Santa Maria (UFSM), Bugwood org
-
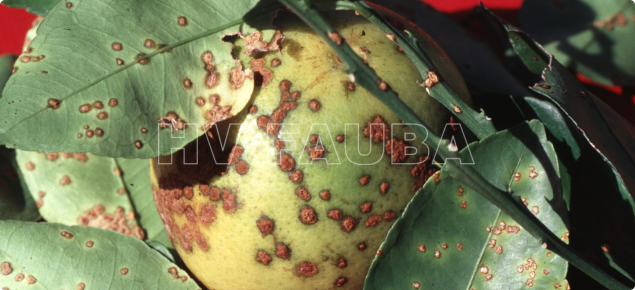
- Autor: Timothy Schubert, Florida Department of Agriculture and Consumer Services, Bugwood org
-

- Autor: M Dewdney
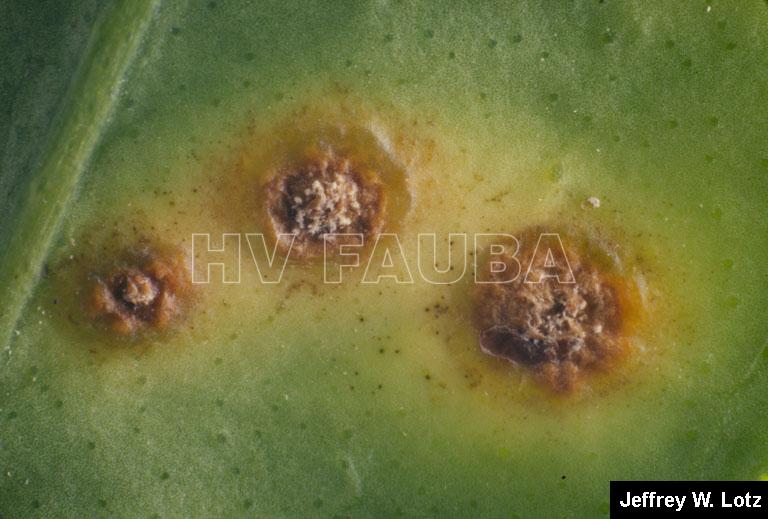
.
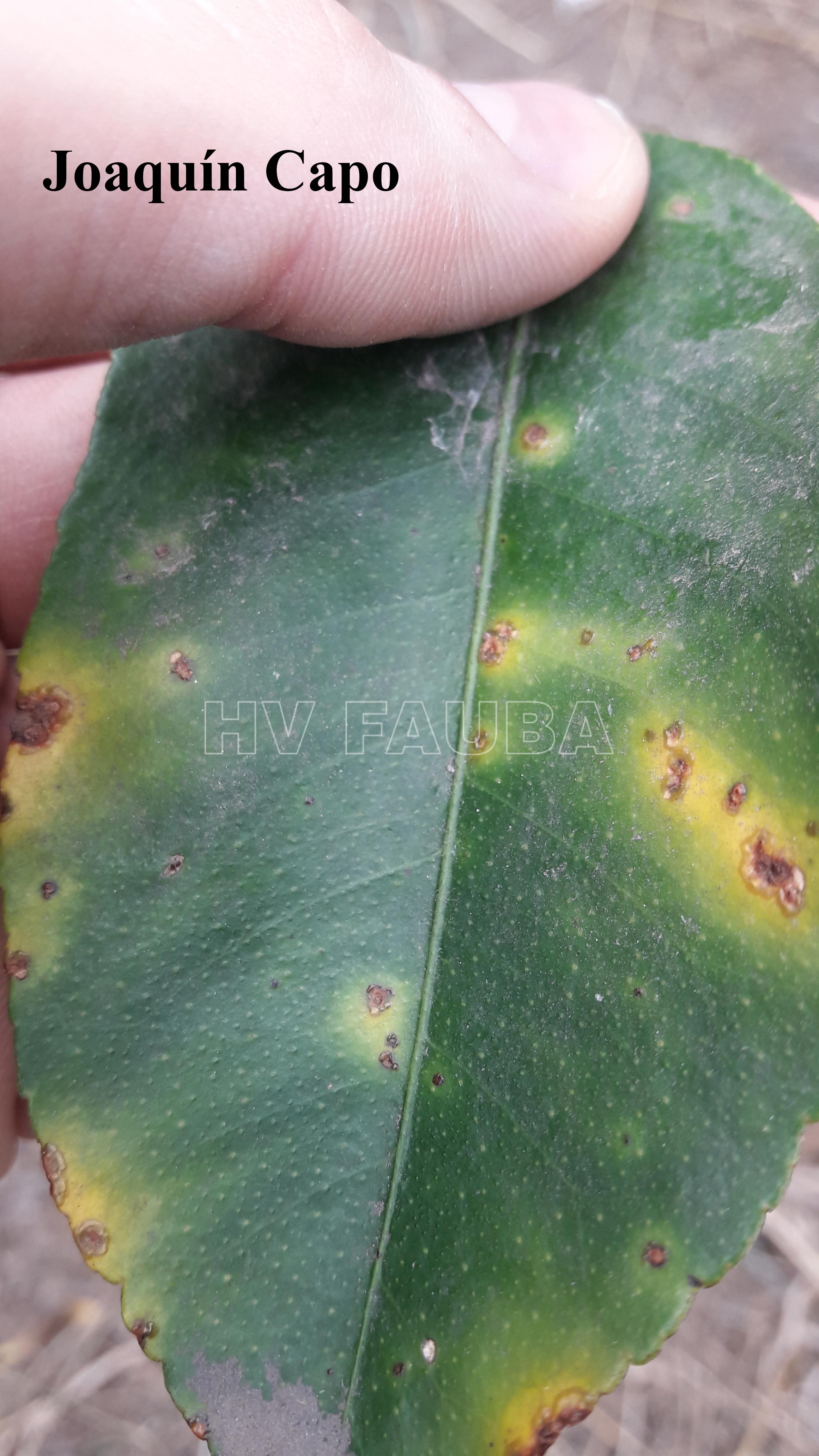
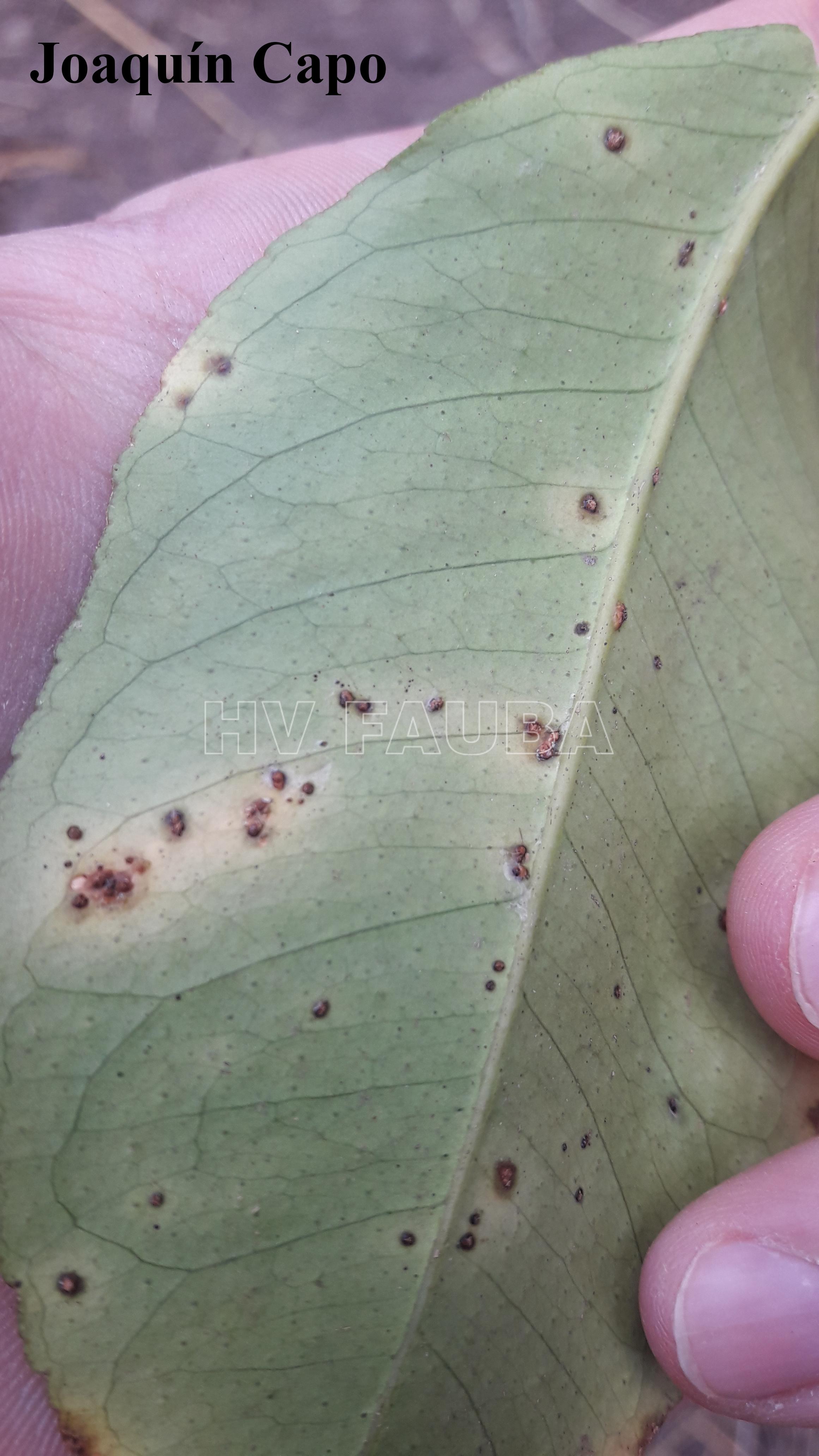
En las hojas se presentan pequeños puntos acuosos traslúcidos o decolorados que se convierten en erupciones de color verde amarillento ubicadas en ambas caras de las hojas; estas erupciones forman una protuberancia circular de tejido suberoso que luego toma una coloración castaño amarillento. Las lesiones están rodeadas por un margen acuoso, donde se localizan las bacterias vivas, seguido de un halo clorótico, también característico de estas lesiones. A medida que las lesiones envejecen adquieren una aspecto corchoso que al rajarse forman una lesión hundida (crateriforme) semejante a un cancro; a este tipo de lesiones se las denomina “pseudocancros”.
En los frutos las lesiones son similares a las de las hojas pero algo más profundas, de color castaño claro y forma irregular, pudiéndose presentar aisladas o confluentes e interesando solamente el epicarpio.
En las ramas las lesiones están constituídas por pequeñas fisuras alargadas y angulares que pueden profundizar y formar verdaderos cancros.
Las pústulas húmedas y los cancros se producen como consecuencia de la inducción del crecimiento celular (hipertrofia) y la división celular (hiperplasia). La inducción ocurre dentro de las 72 h posteriores a la inoculación, como consecuencia de la secreción de la proteína PthA en las células del hospedante. El gen pthA es el responsable de la patogenicidad de Xanthomonas y es necesario para que la bacteria cause la enfermedad (Swarup et al., 1992; Furman, 2013; Li et al., 2014; Ference et al., 2017).
.
-

- Cancrosis en naranjo. Autor: Jonas Janner Hamann, Universidade Federal de Santa Maria (UFSM)

-

- Cancrosis en pomelo. Autor: Timothy Schubert, Florida Department of Agriculture and Consumer Services.







-

- Autor: Dra. Maria Cecilia Perez Piza

-

- Autor: Dra. Maria Cecilia Perez Piza

-

- Autor: Dra. Maria Cecilia Perez Piza

-

- Autor: Dra. Maria Cecilia Perez Piza

-

- Autor: Dra. Maria Cecilia Perez Piza

.
-

- Detalle de las lesiones producidas por Xanthomonas en cítricos. (a) X. axonopodis pv. citrumelo a los 7 d.p.i en el espacio apoplástico de una planta de pomelo. (b) Fotografía tomada con un microscopio electrónico de barrido de un cancro rompiendo la epidermis. (c) Lesión húmeda (water soakin) característica rodeando la pústula o pseudocancro. (d) Pseudocancros rodeados por el halo clorótico amarillento característico de la infección por Xanthomonas. Autores: elaborado por Furman Nicolás, Graham et al., 2004 fotografía (a); Gottwald et al., 2002 fotografías (b), (c), (d).
.
.
Condiciones predisponentes
El principal vector es el hombre ya que promueve su diseminación a larga distancia a través del transporte de plantas completas o partes de plantas, como yemas o varetas, infectadas, movimiento de maquinaria de quintas sanas a infectadas. Hasta el momento no se conoce ninguna epidemia que haya sido ocasionada por una fruta infestada.
Condiciones predisponentes:
* Todo tejido nuevo y en activo crecimiento es susceptible de ser infectado por la bacteria. El tejido joven está presente en hojas y ramas de brotaciones recientes y en los frutos en crecimiento.
* Variedades susceptibles y portainjertos vigorosos.
* Lluvias con vientos fuertes (superiores a 28/km por hora) en primavera y verano favorecen el ingreso de la bacteria a la cavidad subestomática.
* Temperaturas entre 21 y 34ºC.
* Presencia de heridas en hojas y frutos. Las tormentas con vientos fuertes y granizo son factores predisponentes por la generación de heridas (puntos de ingreso de la bacteria).
* Presencia del minador de las hojas de los cítricos que coloca los huevos en el envés de los brotes nuevos: cuando éstos eclosionan, las larvas se alimentan del parénquima esponjoso de las hojas construyendo galerías bajo la epidermis que posibilitan el ingreso de la bacteria.
.
-

- El minador de la hoja de los cítricos, Phyllocnistis citrella, alimentándose de las células epidérmicas debajo de la cutícula; lo que determina una separación de la epidermis al parénquima que está ocupado por el aire. La deformación de las hojas depende del tamaño de la hoja nueva cuando es ocupada por el minador. Más cuando es más joven.
-

- Autor: Dra. Verónica Felipe
-

- Autor: Dra. Verónica Felipe
-

- Autor: Dra. Verónica Felipe
.
.
Ciclo de la enfermedad
En infecciones naturales, los períodos de incubación son generalmente de 5-7 días con algunas excepciones. La duración del período de incubación se rige por varios factores de las bacterias patógenas (especies, modo de entrada, densidad de inóculo y tipo de enfermedad), planta hospedante (especie, edad y órgano) y ambiente (humedad y temperatura). En condiciones climáticas óptimas y con 108 células en un sitio de inoculación artificial, la bacteria causante de la cancrosis de los cítricos (X. citri pv. citri) tarda entre 5 y 7 días en producir el síntoma en las hojas jóvenes y suculentas de una variedad de cítricos susceptible (subaguda). Cuando la infección ocurre tarde en el otoño, las bacterias y el tejido hospedante se vuelven rápidamente latentes debido a las bajas temperaturas. En consecuencia, en ese caso si el período de incubación se extiende hasta la primavera siguiente y, por lo tanto, se produce una infección latente significativa. En inoculaciones artificiales de naranjo dulce (Citrus sinensis) con cepas brasileras, bajo condiciones experimentales controladas, el período medio de incubación en el que el 50% de las plantas mostraron síntomas estuvo fuertemente influenciado por la temperatura y la duración de la humedad de la hoja; donde se registró un período de incubación de 16 y 10 días a 25–35°C, respectivamente; a 12 y 15 °C el período de incubación fue de 30 días para 24 h de duración de mojado foliar (Dalla Pria et al., 2006).
La bacteria permanece viva en los márgenes de las lesiones de hojas y frutos hasta que caen. La bacteria queda a nivel del parénquima en la superficie de los tejidos vegetales. De esta manera, un gran número de factores tales como el agua libre, riego, rocío, lluvias y viento, favorezcan su dispersión. Como resultado, la enfermedad se ha diseminado muy rápidamente por todo el mundo, resultando muy difícil su contención. Sobrevive solamente unos pocos días en el suelo y algunos pocos meses en los restos vegetales incorporados al mismo. Sin embargo puede resistir por años en material herborizado o en restos vegetales que se han conservado secos. La bacteria no se transmite por semilla.
.
-
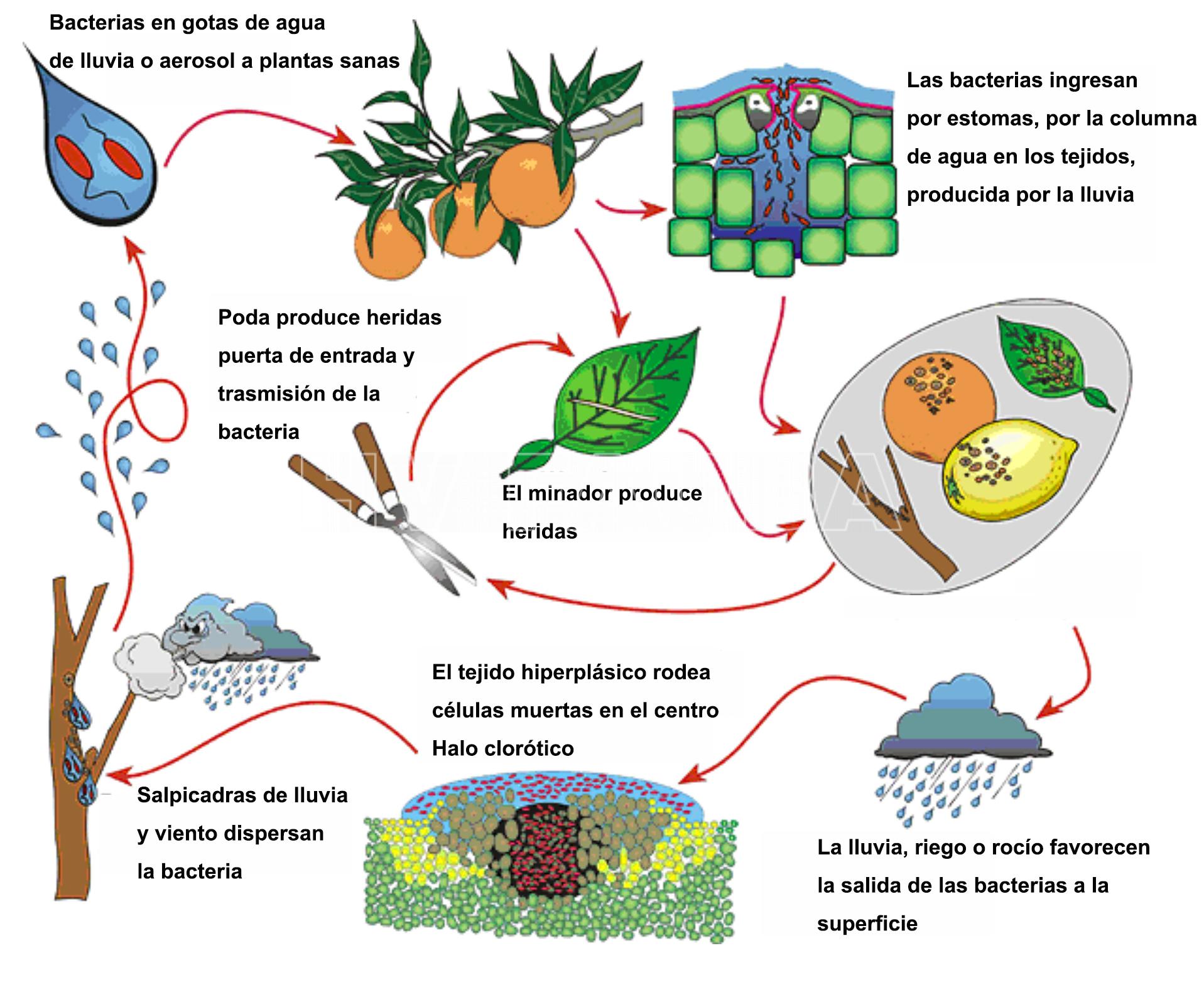
- Fig. 1. Ciclo biológico-agronómico de la Cancrosis de los cítricos (Xanthomonas citri subsp. citri). (Fuente: Gottwald et al., 2002; Traducción: Ana M. Romero)
.
Epidemiología
La incidencia y severidad de la enfermedad después de la introducción variará dependiendo de las condiciones ambientales y de las variedades de cítricos que se cultiven. A partir de las fuentes de inóculo primario (plantas sintomáticas y yemas para injerto infectadas, indumentaria de los operarios y herramientas contaminadas, cajones de madera que han tenido material enfermo, restos de pulpa de fruta procesada, maletas de cosecheros, vehículos que ingresan y transitan en la propiedad, etc.) se produce la primera infección durante la estación de crecimiento, dando lugar a la formación de las lesiones características. Dichas lesiones están embebidas en una sustancia pegajosa que el agua se encarga de diluir; de este modo, la bacteria fluye desde la lesión quedando en las gotitas de agua que son dispersadas por el viento; en este caso, el viento tiene una doble función ya que arrastra la gota de agua acompañada por la bacteria a árboles cercanos a la planta enferma pero, además, provoca una presión en el tejido sano que hace que se forme una columna líquida que permite su paso por el estoma en forma directa dando origen a una nueva lesión. La bacteria penetra por aberturas naturales (estomas, lenticelas) aunque igualmente importantes son las heridas mecánicas producidas por viento, espinas, heladas, incluso las que provoca la larva del minador de la hoja de los cítricos (Phyllocnistis citrella) que, al dejar al descubierto el mesófilo de la hoja, establece una nueva puerta de entrada para la bacteria. No se considera al minador de la hoja como vector de la enfermedad pero sí como un amplificador del nivel de inóculo. Así como el viento y la lluvia cumplen un rol epidemiológico clave en la dispersión de la bacteria (transporte de la misma a corta distancia) el material de injertación está involucrado en el transporte a larga distancia. Las lesiones del año que se generan, como resultado de la primera infección, se convierten en fuentes de inóculo secundario, a partir de las cuales se producen las sucesivas re-infecciones que diseminan la enfermedad en la misma planta y en árboles vecinos, generando una típica enfermedad policíclica (Fig. 1). Las hojas, tallos y frutos se vuelven resistentes a la cancrosis a medida que van madurando. Casi todas las infecciones se producen en hojas y tallos dentro de las 6 semanas después de haber iniciado el crecimiento. El período más crítico para la infección de los frutos es el de los 90 días siguientes a la caída de los pétalos. Cualquier infección posterior resulta en pequeñas lesiones inconspicuas. Las condiciones que retrasan la maduración de los tejidos del hospedante o promueven mayor número de brotaciones, incrementan los riesgos de ataque. En tal sentido, los árboles que vegetan sobre porta injertos menos vigorosos, como el naranjo trifolio, tienden a tener menos cancrosis que aquellos de crecimiento vigoroso, como el limonero rugoso, debido a las diferencias en el número de brotaciones de cada año. Del mismo modo, los árboles jóvenes, generalmente, son más susceptibles que los más viejos.
.
Manejo integrado de la enfermedad
* Uso de material de injertación certificado (adquiridos en viveros inscriptos en INASE)
* Desinfección de guantes y ropa de los cosecheros y de todas las herramientas (tijeras, cajones, escaleras) y maquinarias usadas en la plantación
* Poda sanitaria, selectiva y localizada de ramas, hojas y frutos afectados
* Eliminación de los restos de cosecha
* Pulverizaciones con cobre, teniendo en consideración las brotaciones y las condiciones predisponentes (lluvias y vientos simultáneos)
* Implantar cortinas rompevientos (para disminuir la incidencia del viento)
* Control del minador de los cítricos (químico, biológico)
* Control de malezas
* Evitar fertilizaciones en exceso con nitrógeno
* Usar portainjertos poco vigorosos (P. trifoliata preferible a limón rugoso) (menos brotaciones y menor proporción de tejidos tiernos que presentan susceptibilidad a la bacteria).
* El artículo 19 de la Disposición DNPV N° 4/13 SENASA establece que todo productor de fruta tiene la obligatoriedad de demostrar con la Guía de Sanidad para el Tránsito de plantas y/o sus partes el origen del material de propagación plantado.
.
Desde 1994 hasta 2009 se realizó un importante relevamiento para localizar cepas resistentes a cobre (CuR) en todas las áreas de cultivo de cítricos de Argentina. Se evaluó un total de 3350 cepas, de las cuales aproximadamente el 10% fueron CuR. Las cepas resistentes fueron solo de una región restringida, varias arboledas en el área del río Paraná de la provincia de Corrientes, varias arboledas en la provincia de Formosa en 2004 y pocas arboledas en el SE de Corrientes desde 2007 (Canteros et al., 2013, 2017).
El uso preventivo de la mezcla de mancozeb con cobre al menos una vez al año fue suficiente para detener o retrasar la aparición de cepas de CuR en montes infectados con cepas de CuS. Se recomendó la adición de mancozeb a los productos de cobre inmediatamente después de la primera detección de CuR en Xc-A.
.
Momento de pulverización con productos fitosanitarios (ej. cobre en sus distintos formatos) en el hemisferio sur:
1°) Floración y primera brotación de la temporada (Agosto / Septiembre)
2°) Post-floración, 30-40 días después de la primera pulverización; frutos de 1 cm de diámetro (Septiembre / Octubre)
3°) Tamaño de fruto 2-4 cm (Noviembre / Diciembre); brotación en estado susceptible
4°) Tamaño de fruto 4-5 cm (Diciembre / Enero); brotación en estado susceptible
.
.












-

- Cancrosis en fruto

-

- Síntomas de la Cancrosis de los cítricos en fruto

-

- Lesiones grandes de cancrosis de los cítricos en frutos inmaduros con halos cloróticos. Autor: Gottwald et al., 2002

-

- Autor: M. Goto,
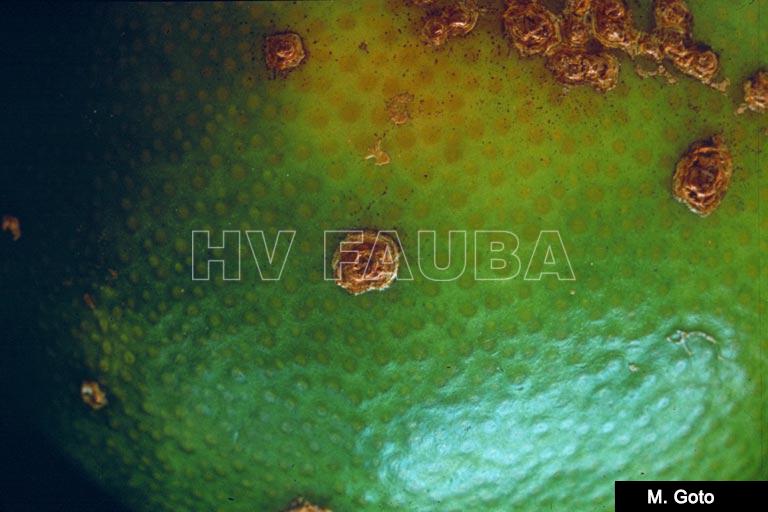
-
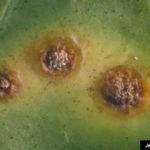
- Autor: Jeffrey W. Lotz, Florida Department of Agriculture and Consumer Services

-

- Autor: Claudio Baron

-

- Autor: Claudio Baron

-

- Autor: Claudio Baron
-

- Autor: Claudio Baron
-

- Autor: Claudio Baron

-

- Autor: Claudio Baron

-

- Autor: Claudio Baron

-

- Autor: Claudio Baron

-

- Autor: Claudio Baron

-

- Autor: Claudio Baron
-

- Autor: Claudio Baron
-

- Autor: Claudio Baron
-

- Autor: Julián Mendiburu
-

- Autor: Julián Mendiburu
-

- De izquierda a derecha: Cancrosis, Sarna, Melanosis. Autor: Claudio Baron
-

- De izquierda a derecha: Cancrosis, Sarna, Melanosis. Autor: Claudio Baron
.
.
Bibliografía
Xanthomonas citri subsp. citri. Sistema Nacional Argentino de Vigilancia y Monitoreo de plagas
Xanthomonas citri pv. citri (XANTCI)[Overview]| EPPO Global Database
Citrus Canker. USDA
Agruirre et al. (2018) Caracterización de aislamientos de Xanthomonas citri pv. citri agente causal de la cancrosis de los cítricos. INTA
Agruirre et al. (2019) Correlación entre patogenicidad y movilidad bacteriana de cepas de Xanthomonas citri subsp. citri. IX Congreso Argentino de Citricultura. INTA
Ahmad AA, Askora A, Kawasaki T, et al. (2014) The filamentous phage XacF1 causes loss of virulence in Xanthomonas axonopodis pv. citri, the causative agent of citrus canker disease. Frontiers in Microbiology 5: 321. doi: 10.3389/fmicb.2014.00321
Alvarez-Martinez CE, Sgro GG, Araujo GG, et al. (2020) Secrete or perish: The role of secretion systems in Xanthomonas biology. Computational and Structural Biotechnology Journal 19: 279-302. doi: 10.1016/j.csbj.2020.12.020
An SQ, Potnis N, Dow M, et al. (2020) Mechanistic insights into host adaptation, virulence and epidemiology of the phytopathogen Xanthomonas. FEMS Microbiology Reviews 44: 1–32. doi: 10.1093/femsre/fuz024
Andrade MO, Zhang Y, Wang W, et al. (2021) Examination of the global regulon of CsrA in Xanthomonas citri subsp. citri using quantitative proteomics and other approaches. Mol Plant Microbe Interact. doi: 10.1094/MPMI-05-21-0113-R
, , , (2023) Suppression of citrus canker disease mediated by flagellin perception. Molecular Plant Pathology 24: 331– 345. doi: 10.1111/mpp.13300
Behlau F, Amorim L, Belasque J, Jr, et al. (2010) Annual and polyetic progression of citrus canker on trees protected with copper sprays. Plant Pathology 59: 1031-1036. doi: 10.1111/j.1365-3059.2010.02344.x
Behlau F, Canteros BI, Minsavage GV, et al. (2011) Molecular characterization of copper resistance genes from Xanthomonas citri subsp. citri and Xanthomonas alfalfae subsp. citrumelonis. Appl Environ Microbiol. 77(12): 4089-4096. doi:10.1128/AEM.03043-10
Behlau F, Gochez AM, Jones JB (2020) Diversity and copper resistance of Xanthomonas affecting citrus. Tropical plant pathology. doi: 10.1007/s40858-020-00340-1
Bejarano SG, Rivadeneira M, Rivadeneira M (2011) Supervivencia de Xanthomonas axonopodis pv citri en suspensiones obtenidas de la superficie de frutos cítricos sin lesiones de cancrosis procesados en empaque. INTA EEA Yuto
Bejarano SG, Rivadeneira M (2011) Comparación de la capacidad de recuperación de Xanthomonas axonopodis pv citri de síntomas de cancrosis de hojas y frutos de cítricos afectados en distintos medios de cultivo semiselectivos. INTA EEA Yuto
Brambilla MV, Piris EB, Mitidieri MS, et al. (2012) Análisis para la detección de Xanthomonas axonopodis pv. citri en fruta cítrica sin síntomas de cancrosis. INTA EEA San Pedro
Campos PE, Groot Crego C, Boyer K, et al. (2021) First historical genome of a crop bacterial pathogen from herbarium specimen: Insights into citrus canker emergence. PLoS Pathog 17(7): e1009714. doi: 10.1371/journal.ppat.1009714
Canteros BI (1999) Copper resistance in Xanthomonas campestris pv. citri. In: Plant pathogenic bacteria, ed. by A. Mahadevan, pp. 455-459. Centre for Advanced Study in Botany, University of Madras, Chennai, India.
Canteros BI, Rybak M, Naranjo M, et al. (2004) Molecular characterization of the copper resistance in Xanthomonas axonopodis pv. citri. In: Book of abstracts. XV. Meeting of Scientific and technical communications of the UNNE.
Canteros BI, Hermosís F, Solíz JA, et al. (2011) Bacteriocins produced by strains of group A
against strains group B of Xanthomonas axonopodis pv. citri citrus canker causing. In: Book of abstracts 2nd Argentine congress of phytopathology. Buenos Aires, Argentina. 184 pp
Canteros BI, Gochez AM, Hermosis F, et al. (2013) Current state of copper resistance situation in the causal bacterium of citrus cancers in Argentina. VII. Argentine Congress of Citriculture, Puerto Iguazu, Misiones. S4. 17 pp.
Canteros BI, Gochez AM, Moschini RC (2017) Management of Citrus Canker in Argentina, a Success Story. The Plant Pathology Journal 33(5): 441-449. doi: 10.5423/PPJ.RW.03.2017.0071
Catara V, Cubero J, Pothier JF, et al. (2021) Trends in Molecular Diagnosis and Diversity Studies for Phytosanitary Regulated Xanthomonas. Microorganisms 9(4): 862. doi: 10.3390/microorganisms9040862
Costa J, Pothier JF, Boch J, et al. (2021) Integrating science on Xanthomonadaceae for sustainable plant disease management in Europe. Mol Plant Pathol. 22(12): 1461-1463. doi: 10.1111/mpp.13150
Dalla Pria M, Christiano RCS, Furtado EL, et al. (2006) Effect of temperature and leaf wetness duration on infection of sweet oranges by Asiatic citrus canker. Plant Pathology 55: 657-663. doi: 10.1111/j.1365-3059.2006.01393.x
Das AK (2003) Citrus canker – A review. J. Appl. Hort.,5(1): 52-60.
de Mello FE, Zaniboni C, Barreto T, et al. (2020) Soil application of acibenzolar-s-methyl and thiamethoxam for the management of citrus canker under subtropical conditions. Trop. plant pathol. 45: 684–690. doi: 10.1007/s40858-020-00367-4
de Souza M, Singh R, Khanal C, et al. (2021) Screening of Commercially Available Satsuma (Citrus unshiu and C. reticulata) Cultivars for Their Susceptibility to Citrus Canker Caused by Xanthomonas axonopodis pv. citri. Plant Health Progress 22: 122-128. doi: 10.1094/PHP-12-20-0105-RS
de Souza-Neto RR, Vasconcelos FNDC, Teper D, et al. (2023) The Expansin Gene CsLIEXP1 Is a Direct Target of CsLOB1 in Citrus. Phytopathology 113(7): 1266-1277. doi: 10.1094/PHYTO-11-22-0424-R
Domingo-Calap ML, Bernabéu-Gimeno M, Aure MC, et al. (2022) Comparative Analysis of Novel Lytic Phages for Biological Control of Phytopathogenic Xanthomonas spp. Microbiol Spectr. e0296022. doi: 10.1128/spectrum.02960-22
Ference CM, Gochez AM, Behlau F, et al. (2018) Recent advances in the understanding of Xanthomonas citri ssp. citri pathogenesis and citrus canker disease management. Molecular Plant Pathology 19: 1302-1318. doi: 10.1111/mpp.12638
Ferreira DH, Moreira RR, Silva Junior GJ, et al. (2022) Copper rate and spray interval for joint management of citrus canker and citrus black spot in orange orchards. Eur J Plant Pathol. doi: 10.1007/s10658-022-02527-5
Fonseca NP, Patané JSL, Varani AM, et al. (2019) Analyses of Seven New Genomes of Xanthomonas citri pv. aurantifolii Strains, Causative Agents of Citrus Canker B and C, Show a Reduced Repertoire of Pathogenicity-Related Genes. Front. Microbiol. 10: 2361. doi: 10.3389/fmicb.2019.02361
Furman F (2013) Desarrollo de alternativas biotecnológicas para la obtención de plantas de Citrus sinensis (L.) Osbeck resistentes a la enfermedad de la Cancrosis de los cítricos. Tesis Doctoral, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. 156 p.
Garita-Cambronero J, Sena-Vélez M, Ferragud E, et al. (2019) Xanthomonas citri subsp. citri and Xanthomonas arboricola pv. pruni: Comparative analysis of two pathogens producing similar symptoms in different host plants. PLoS ONE 14(7): e0219797. doi: 10.1371/journal.pone.0219797
Gochez AM, Shantharaj D, Potnis N, et al. (2017) Molecular characterization of XopAG effector AvrGf2 from Xanthomonas fuscans ssp. aurantifolii in grapefruit. Molecular Plant Pathology 18: 405–419. doi: 10.1111/mpp.12408
Gochez AM, Behlau F, Singh R, et al. (2020) Panorama of citrus canker in the United States. Tropical Plant Pathology 45: 192–199. doi: 10.1007/s40858-020-00355-8
Gonçalves-Zuliani AMO, et al. (2016) Reaction of detached leaves from different varieties of sweet orange to inoculation with Xanthomonas citri subsp. citri. Summa Phytopathologica 42(2): 125-133. doi: 10.1590/0100-5405/2143
Gottig N, Vranych CV, Sgro GG, et al. (2018) HrpE, the major component of the Xanthomonas type three protein secretion pilus, elicits plant immunity responses. Scientific Reports 8: 9842. doi: 10.1038/s41598-018-27869-1
Gottwald TR, Graham JH, Schubert TS (2002) Citrus canker: The pathogen and its impact. Online. Plant Health Progress doi: 10.1094/PHP-2002-0812-01-RV
Graham JH, Johnson EG, Myers ME, et al. (2016) Potential of Nano-Formulated Zinc Oxide for Control of Citrus Canker on Grapefruit Trees. Plant Disease 100(12): 2442-2447. doi: 10.1094/PDIS-05-16-0598-RE
Haq F, Xu X, Ma W, et al. (2021) A Xanthomonas transcription activator-like effector is trapped in nonhost plants for immunity. Plant Communications. doi: 10.1016/j.xplc.2021.100249
Huang R, Hui S, Zhang M, et al. (2017) A Conserved Basal Transcription Factor Is Required for the Function of Diverse TAL Effectors in Multiple Plant Hosts. Frontiers in Plant Science 8: 1919. doi: 10.3389/fpls.2017.01919
Huang F, Ling J, Zhu C, et al. (2023) Canker Disease Intensifies Cross-Kingdom Microbial Interactions in the Endophytic Microbiota of Citrus Phyllosphere. Phytobiomes Journal. doi: 10.1094/PBIOMES-11-22-0091-R
Islam MN, Ali MS, Choi SJ, et al. (2019)Biocontrol of Citrus Canker Disease Caused by Xanthomonas citri subsp. citri Using an Endophytic Bacillus thuringiensis. The Plant Pathology Journal 35(5): 486-497. doi: 10.5423/PPJ.OA.03.2019.0060
Izadiyan M, Taghavi SM (2020) Isolation and characterization of the citrus canker pathogen Xanthomonas citri subsp. citri pathotype A, occurring in imported tangerine (Citrus reticulata Blanco) fruits. J Plant Pathol 102: 671–679. doi: 10.1007/s42161-020-00568-5
Juhasz CC, Leduc A, Boyer C, et al. (2013) First Report of Xanthomonas citri pv. citri Causing Asiatic Citrus Canker in Burkina Faso. Plant Disease 97(12): 1653. doi: 10.1094/PDIS-06-13-0600-PDN
Li N, Huang L, Liu L, et al. (2014) The relationship between PthA expression and the pathogenicity of Xanthomonas axonopodis pv. citri . Mol Biol Rep 41: 967–975. doi: 10.1007/s11033-013-2941-4
Li W, Song Q, Brlansky RH, Hartung JS (2007) Genetic diversity of citrus bacterial canker pathogens preserved in herbarium specimens. PNAS 104 (47): 18427-18432. doi: 10.1073/pnas.0705590104
Lorenzoni ASG, Dantas GC, Bergsma T, et al. (2017) Xanthomonas citri MinC Oscillates from Pole to Pole to Ensure Proper Cell Division and Shape. Front. Microbiol. 8: 1352. doi: 10.3389/fmicb.2017.01352
Malamud F (2011) Bases moleculares del desarrollo de biofilms en Xanthomonas axonopodis pv citri y su rol en el proceso infectivo. Tesis Doctoral, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. 135 p.
Martínez V, Acosta E, Rojas A, et al. (2014) Estudios de sobrevivencia de Xanthomonas citri subsp. citri en frutos de limoneros y su dispersión a partir de frutos cosechados y severamente infectados. Revista industrial y agrícola de Tucumán 91(2): 17-24. versión On-line ISSN 1851-3018
Martins PMM, de Oliveira Andrade M, Benedetti CE, et al. (2020) Xanthomonas citri subsp. citri: host interaction and control strategies. Tropical Plant Pathology 45: 213–236. doi: 10.1007/s40858-020-00376-3
Melotto M, Underwood W, He SY (2008) Role of stomata in plant innate immunity and foliar bacterial diseases. Annu Rev Phytopathol. 46: 101-22. doi: 10.1146/annurev.phyto.121107.104959
Michalopoulou VA, Vicente JG, Studholme DJ, Sarris PF (2018) Draft Genome Sequences of Pathotype Strains for Three Pathovars Belonging to Three Xanthomonas Species. Microbiology Resource Announcement 7(12): e00923-18. doi: 10.1128/MRA.00923-18
, , , , The Arabidopsis immune receptor EFR increases resistance to the bacterial pathogens Xanthomonas and Xylella in transgenic sweet orange.
Moschini RC, Canteros BI, Martínez MI, et al. (2014) Quantification of the environmental effect on citrus canker intensity at increasing distances from a natural windbreak in northeastern Argentina. Australasian Plant Pathology 43: 653–662. doi: 10.1007/s13313-014-0305-8
Nazaré AC, Polaquini CR, Cavalca LB, et al. (2018) Design of Antibacterial Agents: Alkyl Dihydroxybenzoates against Xanthomonascitri subsp. citri. Int J Mol Sci. 19(10): 3050. doi: 10.3390/ijms19103050
Patané JSL, Martins J, Rangel LT, et al. (2019) Origin and diversification of Xanthomonas citri subsp. citri pathotypes revealed by inclusive phylogenomic, dating, and biogeographic analyses. BMC Genomics 20: 700. doi: 10.1186/s12864-019-6007-4
Perez E, Kunta M, Ancona V, et al. (2021) The return of asiatic citrus canker to texas: surveys and eradication efforts. Plant Health Progress. doi: 10.1094/PHP-10-20-0089-S
Rojas AA, Salas H, Fogliata GM, Martínez CV (2011) Eficacia de ingredientes activos cúpricos para el control de la cancrosis de los cítricos en limón, Tucumán, 2008/09 y 2009/10. 2º Congreso Argentino de Fitopatología. Libro de resúmenes. p. 266.
Rossini MN, Dummel DM, Agostini JP (2015) Plagas cuarentenarias de frutales de la República Argentina. INTA. Link
Russi P, Menoni M, del Campo R, Peyrou M (2013) Caracterización de cepas de Xanthomonas citri sbsp. citri, agente causal del cancro cítrico. Agrociencia Uruguay 17( 2 ): 64-74. versión On-line ISSN 2301-1548
Ryan RP, Vorhölter FJ, Potnis N, et al. (2011) Pathogenomics of Xanthomonas: understanding bacterium-plant interactions. Nature Reviews Microbiology 9(5): 344–355. doi: 10.1038/nrmicro2558
Schubert TS, Rizvi SA, Sun X, et al. (2001) Meeting the challenge of Eradicating Citrus Canker in Florida—Again. Plant Disease 85(4): 340-356. doi: 10.1094/PDIS.2001.85.4.340
Sgro GG, Ficarra FA, Dunger G, et al. (2012) Contribution of a harpin protein from Xanthomonas axonopodis pv. citri to pathogen virulence. Molecular Plant Pathology 13: 1047–1059. doi: 10.1111/j.1364-3703.2012.00814.x
Silva IC, Regasini LO, Petrônio MS, et al. (2013) Antibacterial activity of alkyl gallates against Xanthomonas citri subsp. citri. J Bacteriol. 195(1): 85-94. doi: 10.1128/JB.01442-12
Soares JM, Tanwir SE, Grosser JW, et al. (2020) Development of genetically modified citrus plants for the control of citrus canker and huanglongbing. Tropical Plant Pathology 45: 237–250. doi: 10.1007/s40858-020-00362-9
Stall RE, Marco GM, Canteros BI (2015) Cancrosis de los citrus: Proyecto cooperativo INTA – IFAS. – 2da. Edición. — Publicación EEA Bella Vista. Serie Técnica Nº 54. 2015. 57 pp.
Swarup S, Yang Y, Kingsley MT, Gabriel DW (1992) An Xanthomonas citri pathogenicity gene, pthA, pleiotropically encodes gratuitous avirulence on nonhosts. Mol Plant Microbe Interact. 5(3): 204-13. doi: 10.1094/mpmi-5-204
Taylor EL, Gottwald TR, Adkins S (2023) Structural changes in Florida citrus production, 1980-2021 and associated consequences of weather events and disease. Journal of Citrus Pathology, 10. doi: 10.5070/
Teper D, White FF, Wang N (2023) The Dynamic Transcription Activator-Like Effector Family of Xanthomonas. Phytopathology 113(4): 651-666. doi: 10.1094/PHYTO-10-22-0365-KD
Trinh J, Li T, Franco JY, et al. (2022) Citrus relatives exhibit natural variation in perception and response magnitude to microbial features. bioRxiv 2022.07.15.500235; doi: 10.1101/2022.07.15.500235
Velazquez PD, Farías MF, Famá SA (2007) Relación entre el momento de control químico del minador de la hoja de los cítricos (Phyllocnistis citrella) y la intensidad de cancrosis en brotes de limón (Citrus limon) no tratados con bactericidas cúpricos. Conference: XXX Congreso Argentino de Horticultura y I Simposio Internacional sobre Cultivos Protegidos, 25-28 de septiembre. Resumen FR SV 025. p. 215. ISBN 978-987-97812-1-0., At La Plata, Buenos Aires, Argentina
Vieira PS, Bonfim IM, Araujo EA, et al. (2021) Xyloglucan processing machinery in Xanthomonas pathogens and its role in the transcriptional activation of virulence factors. Nat Commun 12: 4049. doi: 10.1038/s41467-021-24277-4
Wang W, Feng M, Li X, et al. (2022) Antibacterial Activity of Aureonuclemycin Produced by Streptomyces aureus Strain SPRI-371. Molecules 27(15): 5041. doi: 10.3390/molecules27155041
Yang LC, Gan YL, Yang LY, et al. (2018) Peptidoglycan hydrolysis mediated by the amidase AmiC and its LytM activator NlpD is critical for cell separation and virulence in the phytopathogen Xanthomonas campestris. Molecular Plant Pathology 19(7): 1705-1718. doi: 10.1111/mpp.12653
Zekri M, Dewdney M, Burrow J, Roberts P (2015) Identificación en campo de síntomas de cancro de los cítricos y los procedimientos de descontaminación. PP-214-SP, Plant Pathology Department, UF/IFAS Extensión