.
Condición fitosanitaria: Presente ampliamente distribuida
Grupo de cultivos: Cereales
Rango de hospedantes: específico / estrecho
Hospedante (primario): Maíz (Zea mays) (estadios telial/uredinial)
Hospedante intermediario: Oxalis conorrhiza Jacq. (syn. O. cordobensis R. Knuth) (estadios pycnial / aecial) (Guerra et al., 2016)
Etiología: policíclica, subaguda
Etiología: Hongo. Basidiomycota. Biotrófico
Agente causal: Puccinia sorghi Schwein
Taxonomía: Eukaryota > Fungi > Dikarya > Basidiomycota > Pucciniomycotina > Pucciniomycetes > Pucciniales > Pucciniaceae > Puccinia
.
-
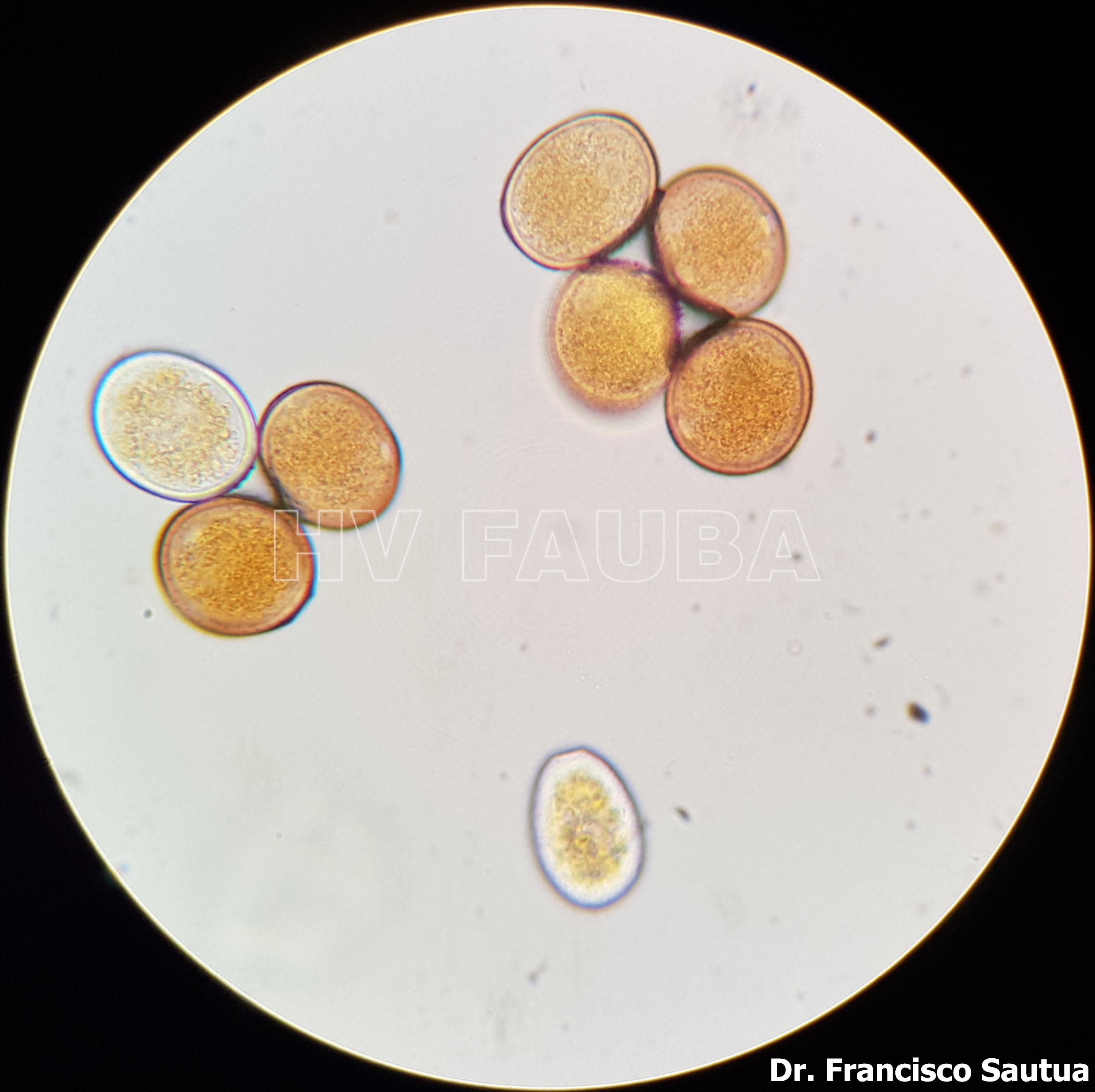
- Uredosporas de Puccinia sorghi al microscopio óptico. Autor: Dr. Francisco Sautua
.
.
.
Síntomas y signos
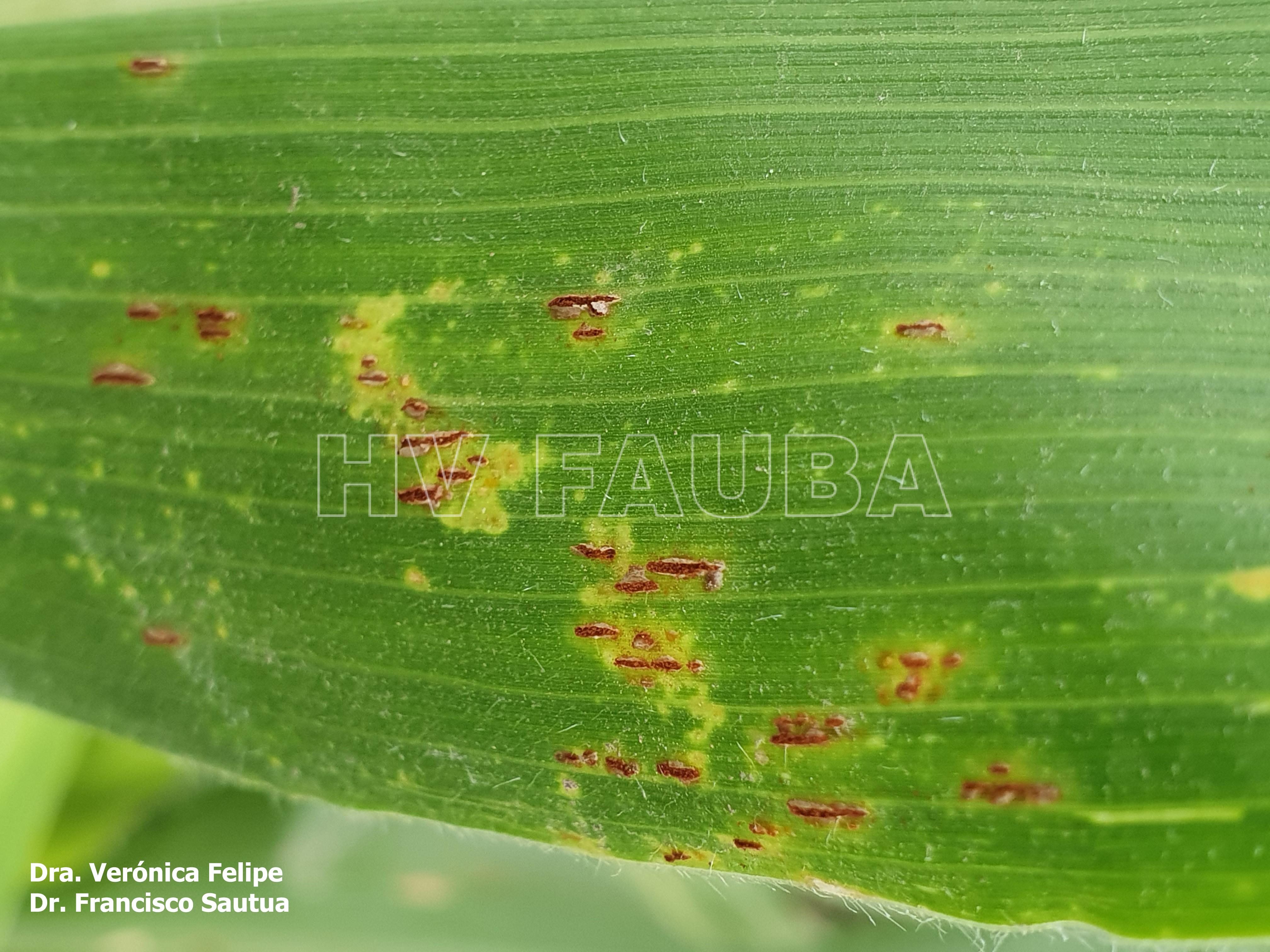
Los primeros síntomas en manifestarse son puntos cloróticos en la superficie de la hoja. Rápidamente se desarrollan, en cultivares susceptibles, en pústulas grandes, circulares a alargadas, pulverulentas, presentando el interior de coloración pardo-canela, luego de romper la epidermis de la hoja. Estas pústulas (uredias) se manifiestan en cualquier tejido verde de la planta. Se encuentran con frecuencia sobre las dos superficies de la hoja. El tejido foliar que rodea las pústulas puede devenir amarillo clorótico o puede morir, dejando pequeñas o grandes áreas de lesiones. Luego las pústulas se vuelven negras, cuando los uredosporas son substituidos por las teliosporas, las cuales continúan su erupción rompiendo la epidermis, a medida que la planta madura.
.
-

- Roya común del maíz, causada por Puccinia sorghi. (a – c) Esporulación sobre híbridos susceptibles, (d) síntomas (clorosis) causados por la acción de parasitismo del patógeno, (e) detalle de pústula uredosórica, (f) detalle de pústula uredosórica con lupa, (g) detalle de uredosporas al miscroscopio óptico. Autores: Jonathan Damiani (a, b, c), Verónica Felipe y Francisco Sautua (d), Francisco Sautua (e, f, g). Autor: Carmona y Sautua, 2021.
.
Antecedentes y Daños
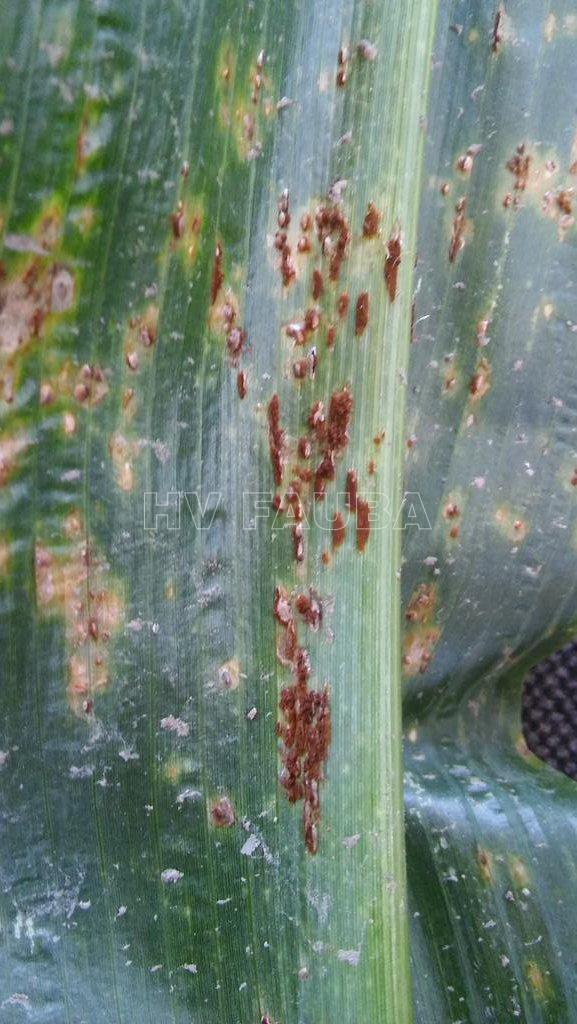
Esta enfermedad es endémica en la zona núcleo (González M., 2005; Sillon et al., 2007). También se encuentra difundida en el NOA (Diaz et al., 2007) y Entre Ríos (Formento, 2010). Este patógeno disminuye el IAFS. En ataques severos las pústulas formadas pueden provocar la necrosis del tejido foliar dando un aspecto de mancha foliar (Figura 29), que puede dificultar el diagnóstico. Hojas enteras pueden morir si son infectadas severamente. Esta roya es más notable hacia floración. En el campo presenta diferentes grados de intensidad cada año de acuerdo al comportamiento de los híbridos utilizados. El ambiente es más predisponente cuando se combinan siembras tardias y escasez de lluvias. Los daños registrados en el mundo oscilan entre un 10 al 30%. En Argentina la presencia de la roya en maíz es frecuente aunque los ataques severos son esporádicos. En determinados años favorables, los daños y pérdidas agronómicas pueden ser elevados. Un hecho para destacar es que durante las últimas campañas se ha observado un importante crecimiento de la roya sobre varios campos de maíz. Durante la campaña 2007-2008 ocurrió una importante epifitia en la principal zona maicera del país desde estadios vegetativos, que obligó al uso de fungicidas.
.
-

- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua
.
Epidemiología
La roya común del maíz ataca solamente al maíz, a pesar de que su nombre haga referencia al sorgo. En Argentina este hongo biotròfico cumple su ciclo completo, generando alta variabilidad genética, comprobándose al menos la existencia de cuatro razas distintas. El agente etiológico es un hongo heteroico y macrocíclico que cumple aparentemente su ciclo en Oxalis spp. (estado picnídico y ecídico), y la otra parte (uredosórico y teleutosórico) en maíz. La detección de picnios y ecidios en Oxalis se ha informado en Argentina sólo en 1952, desde ese año no hay publicaciones al respecto y en monitoreos actuales sólo se ha encontrado pústulas uredosoricas de Puccinia oxalidis que es la roya propia del Oxalis. Las teliosporas provenientes del maíz pueden germinar formando basidiosporas que no son infectivas para el maíz pero si pueden infectar al Oxalis. En este último hospedante intermediario se forman las picniosporas y ecidiosporas. Estas últimas son infectivas al maíz generando uredosporas (Lindquist, 1982). Una vez infectado el maíz (infección proveniente del inoculo primario –ecidiosporas-del Oxalis o por uredosporas traídas por el viento), desde una hoja, planta o campo se generan más uredosporas para infectar nuevas hojas, plantas o campos. Las uredosporas pueden sobrevivir en la naturaleza en plantas guachas que se comportan también como fuentes de inóculo primario.
.
-

- Autor: Tom Allen
.
Supervivencia
Para establecer experimentalmente la relación entre las formas de la fase gametófita encontrada sobre O. corniculata y las correspondientes a la fase esporófita sobre maíz, Godoy y Bruni (1952) durante 1948 y 1949 trabajando en invernáculo en Pergamino inocularon plantitas de maíz cultivadas en maceta, con ecidiosporos desarrollados naturalmente sobre Oxalis corniculata L. (clasificados por el Instituto de Botánica del Ministerio de Agricultura). A los 8 -12 días de la inoculación aparecieron las pústulas uredospódicas, y aproximadamente 30 días más tarde, se formaron los teleutosoros. A la inversa, plantitas de O. corniculata obtenidas de semillas (plantas sanas), fueron inoculadas con teleutosporas presentes en hojas secas de maíz recogidas en el invierno. Después de un período de incubación de 8-9 días se registró la aparición de los primeros picnios y 4 días después, en estrecha relación con las colonias de picnios se formaron los ecidios. Godoy y Bruni (op. cit.) observaron que O. articulata L., muy común en la zona de Pergamino, se mostró inmune al ser inoculado con teleutosporos de P. sorghi. Ambas especies de Oxalis son muy semejantes pero difieren en el color de las flores, amarillas en el caso de corniculata y rosadas o violáceas en el caso de articulata (Antonelli, 2012).
En Argentina se ha confirmado a Oxalis conorrhiza como hospedante intermediario y fuente de inóculo y supervivencia (Guerra et al., 2016). Las plantas guachas de maíz jugarían un rol muy importante en la supervivencia de este patógeno.
.
-
- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua
.
Condiciones predisponentes
Susceptibilidad de los genotipos y abundancia del inóculo. Los tejidos más jóvenes o estados juveniles son más susceptibles. La temperatura ideal para germinación de esporas es 17ºC, aunque buena germinación ocurre entre 13 a 27ºC. La formación de una pústula puede llevar entre 5 a 7 días a 20-25ºC. Si el ataque de roya es severo incrementa las pudriciones de raíz y tallo. La germinación de esporas se muy baja con temperaturas > a 32ºC (Cina et al., 2009). A veces se asocian epidemias con siembras tardías y escasez de lluvia.
.
-

- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua
.
Manejo integardo de la enfermedad
* Siembra de híbridos resistentes/tolerantes
* Uso de fungicidas de acuerdo con el UDE.
.
.
-

- 01 Roya común del maíz, causada por P. sorghi. Híbrido DK6910 en V6 (siembra 25/11/2017), Córdoba. Autor: Ing. Agr. Juan Pablo Ioele.

-

- 02 Roya común del maíz, causada por P. sorghi. Híbrido DK6910 en V6 (siembra 25/11/2017), Córdoba. Autor: Ing. Agr. Juan Pablo Ioele.

-

- 03 Roya común del maíz, causada por P. sorghi. Híbrido DK6910 en V6 (siembra 25/11/2017), Córdoba. Autor: Ing. Agr. Juan Pablo Ioele.

-

- 04 Roya común del maíz, causada por P. sorghi. Híbrido DK6910 en V6 (siembra 25/11/2017), Córdoba. Autor: Ing. Agr. Juan Pablo Ioele.
-

- 05 Roya común del maíz, causada por P. sorghi. Híbrido DK6910 en V6 (siembra 25/11/2017), Córdoba. Autor: Ing. Agr. Juan Pablo Ioele.

-

- 06 Híbridos de maíz atacados por Roya común del maiz, causada por P. sorghi. Córdoba. Autor: Ing. Agr. Juan Pablo Ioele.

-

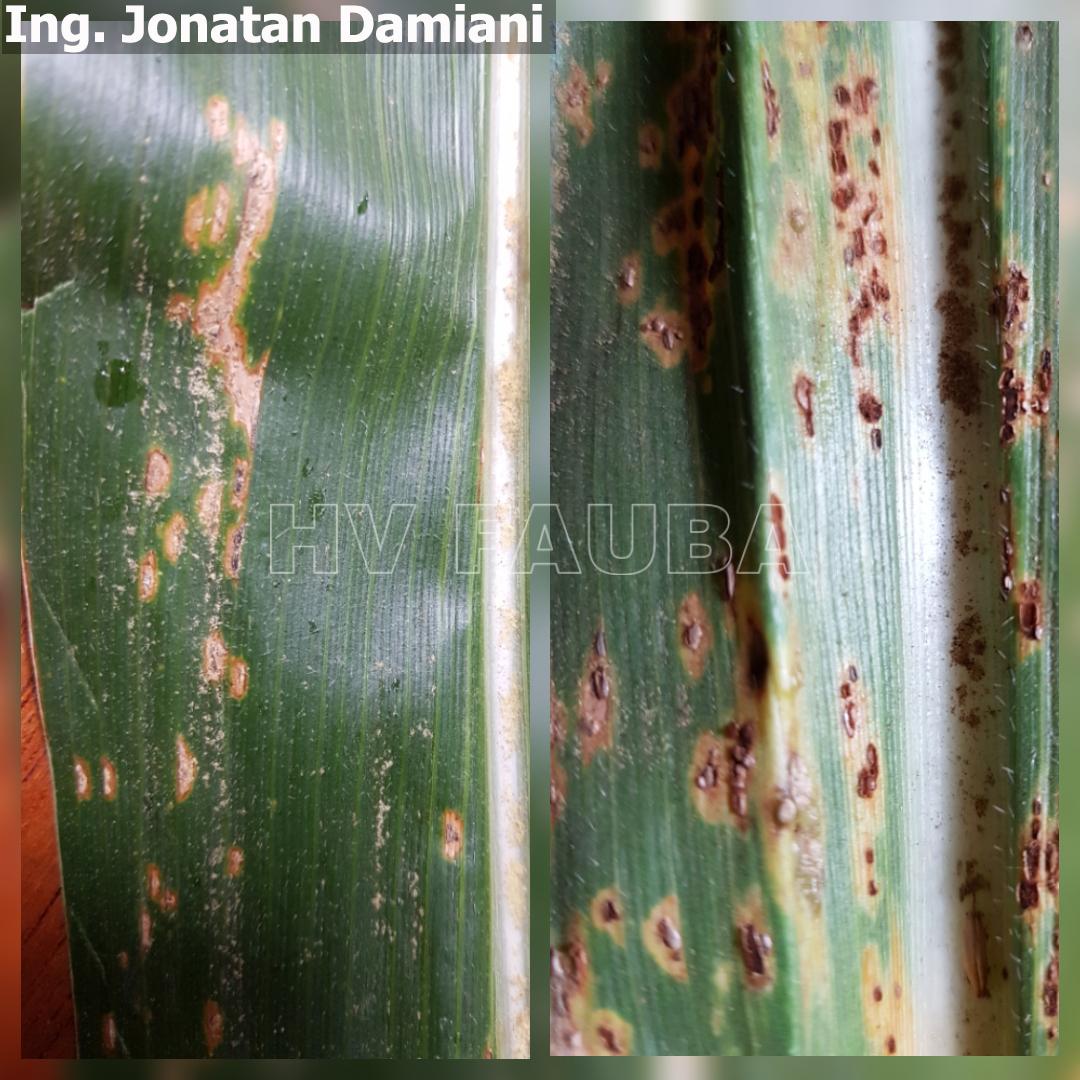
- 01 Síntomas y signo (pústulas uredosóricas) de roya común del maíz, causada por Puccinia sorghi. Híbrido DK 6910 VT3 RIB, Fecha siembra 10/12/17, Sancti Spiritu, Santa Fe. Autor: Ing. Jonatan Damiani

-

- 02 Síntomas y signo (pústulas uredosóricas) de roya común del maíz, causada por Puccinia sorghi. Híbrido DK 6910 VT3 RIB, Fecha siembra 10/12/17, Sancti Spiritu, Santa Fe. Autor: Ing. Jonatan Damiani

-

- 03 Síntomas y signo (pústulas uredosóricas) de roya común del maíz, causada por Puccinia sorghi. Híbrido DK 6910 VT3 RIB, Fecha siembra 10/12/17, Sancti Spiritu, Santa Fe. Autor: Ing. Jonatan Damiani

-

- 04 Control químico de roya común del maíz, causada por Puccinia sorghi, Maíz Tardío, Híbrido DK 6910 VT3 RIB, Fecha siembra 10/12/17, Sancti Spiritu, Santa Fe. Aplicaciones de Fungicidas (estrobilurina + triazol) en V12/13 (08/02/2018). Autor: Ing. Jonatan Damiani

-

- 05 Control químico de roya común del maíz, causada por Puccinia sorghi, Maíz Tardío, Híbrido DK 6910 VT3 RIB, Fecha siembra 10/12/17, Sancti Spiritu, Santa Fe. Aplicaciones de Fungicidas (estrobilurina + triazol) en V12/13 (08/02/2018). Autor: Ing. Jonatan Damiani

-

- 06 Control químico de roya común del maíz, causada por Puccinia sorghi, Maíz Tardío, Híbrido DK 6910 VT3 RIB, Fecha siembra 10/12/17, Sancti Spiritu, Santa Fe. Aplicaciones de Fungicidas (estrobilurina + triazol) en V12/13 (08/02/2018). Autor: Ing. Jonatan Damiani




-

- Sintomas y Signos de la Roya comón del maíz causada por Puccinia sorghi. Autor: Dirceu Gassen

-

- Sintomas y Signos de la Roya comón del maíz causada por Puccinia sorghi. Autor: Dirceu Gassen

-

- Pustulas uredosórica de roya común del maíz (P. sorghi). Autor: Dr. Marcelo Carmona

-

- Pustulas uredosórica de roya común del maíz (P. sorghi). Autor: Dr. Marcelo Carmona

-

- Pustula uredosórica de roya común del maíz (P. sorghi). Autor: Dr. Marcelo Carmona

-

- Pústulas uredosóricas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-

- Pústulas uredosóricas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-

- Pústulas uredosóricas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-

- Pústulas uredosóricas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-

- Pústulas uredosóricas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-

- Pústulas uredosóricas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-

- Pústulas uredosóricas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-
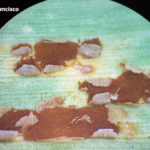
- Pústulas uredosóricas con uredosporas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-

- Pústulas uredosóricas con uredosporas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-
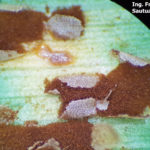
- Pústulas uredosóricas de la Roya Común del maíz, causada por Puccinia sorghi. Autor: Dr. Francisco Sautua

-
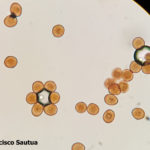
- Uredosporas de Puccinia sorghi. Autor: Dr. Francisco Sautua
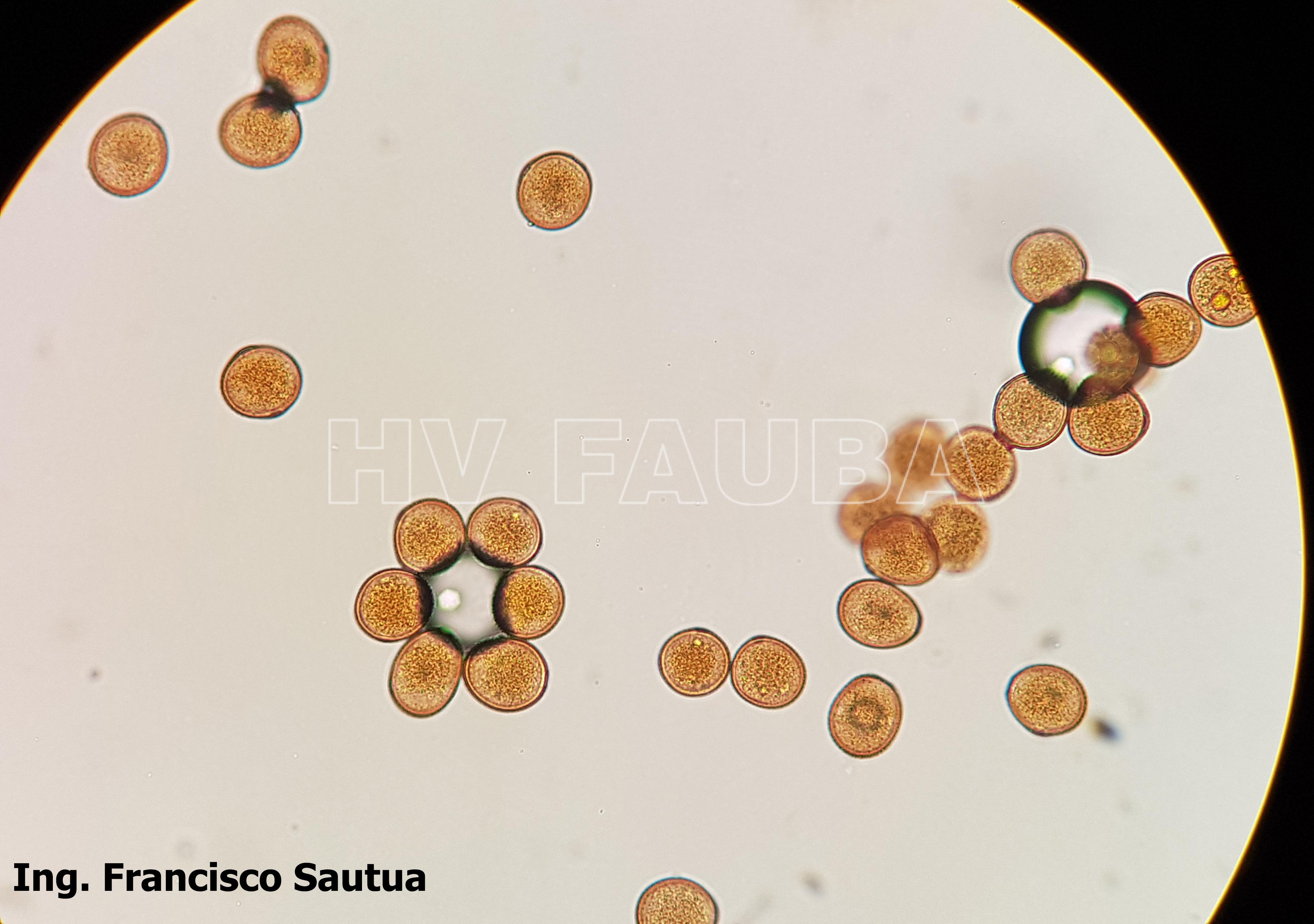
-
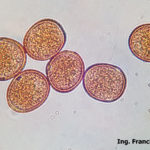
- Uredosporas de Puccinia sorghi. Autor: Dr. Francisco Sautua
-
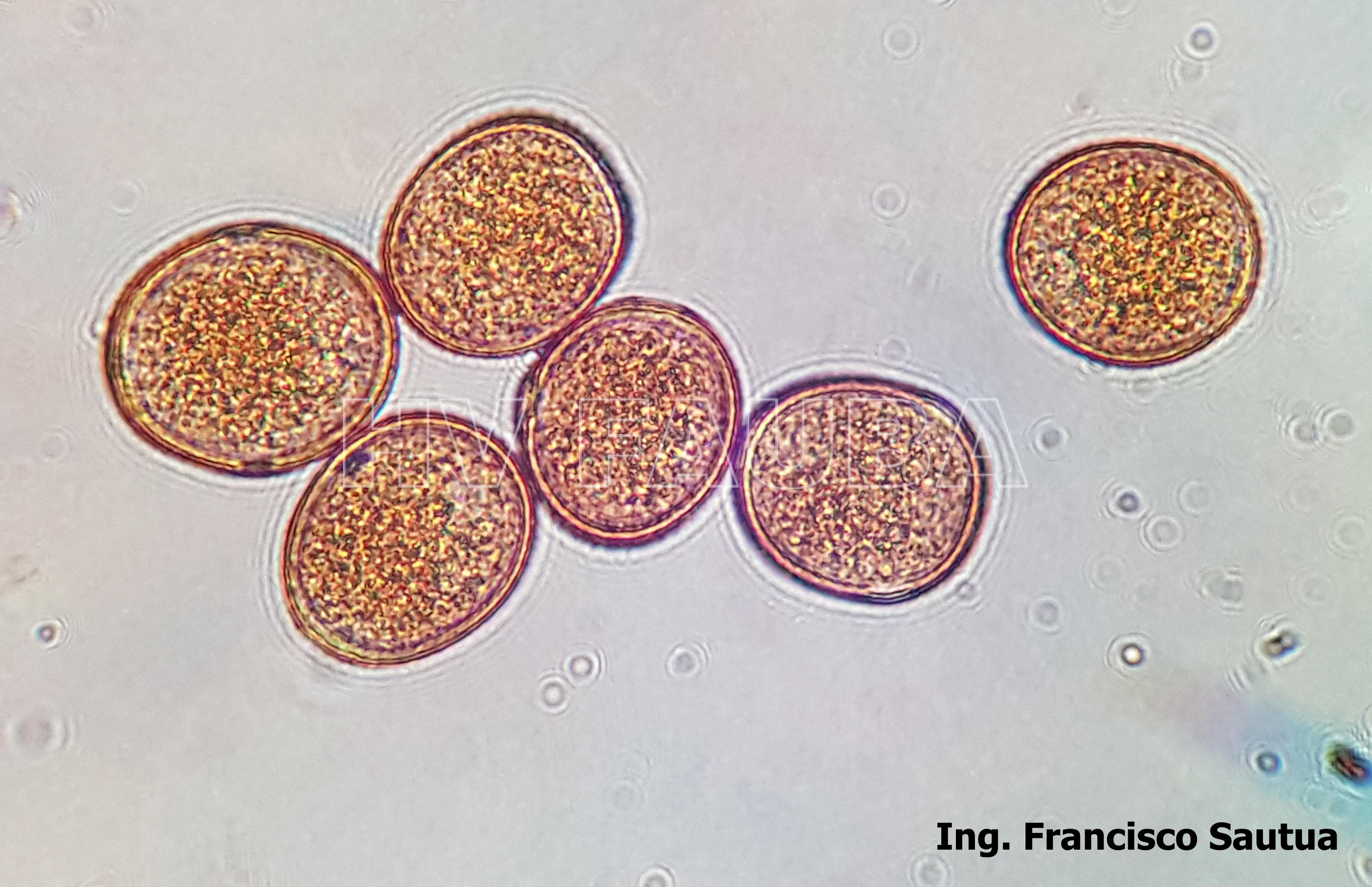
- Uredosporas de Puccinia sorghi. Autor: Dr. Francisco Sautua
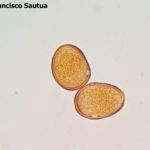
-

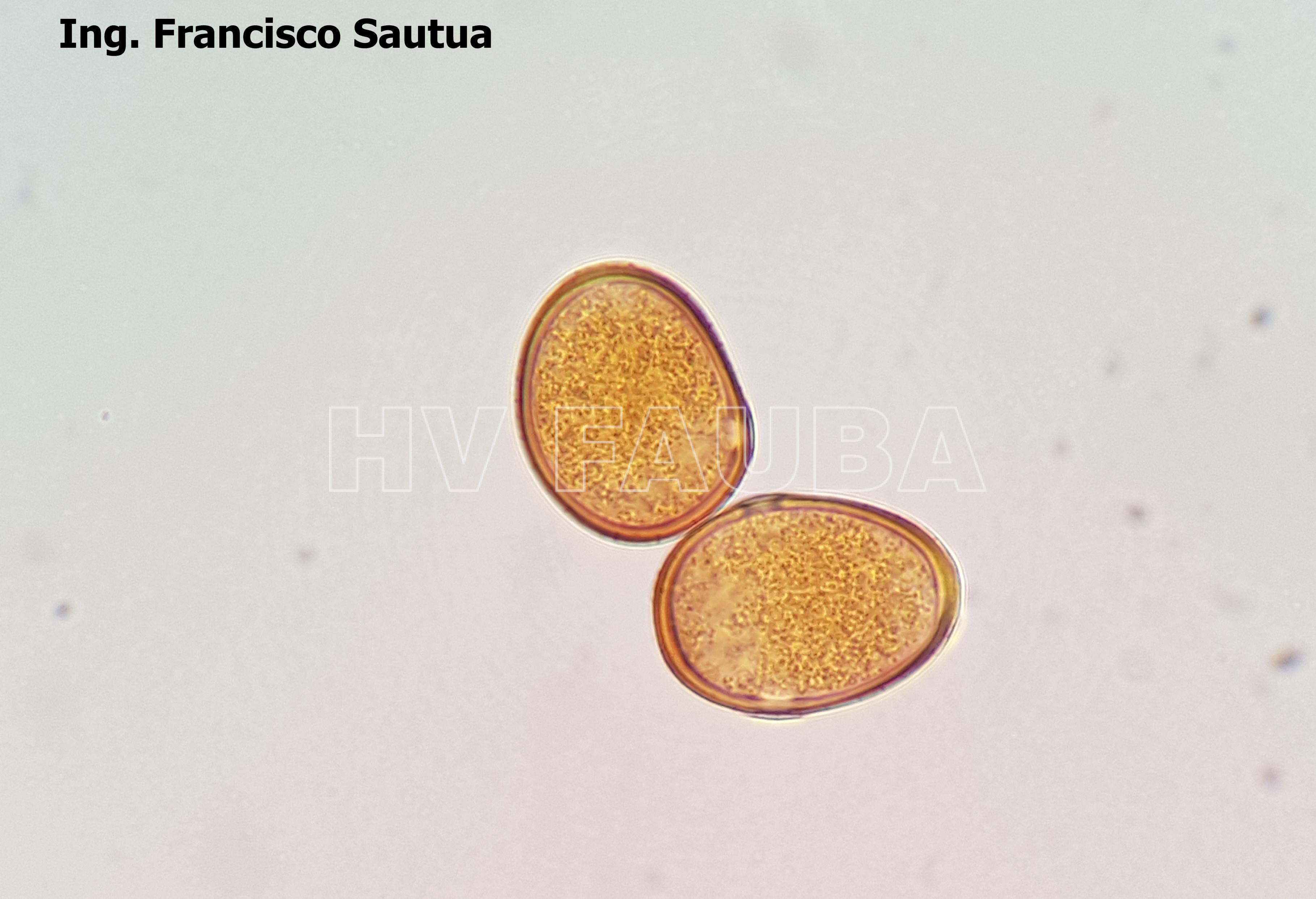
- Uredosporas de Puccinia sorghi. Autor: Dr. Francisco Sautua

-
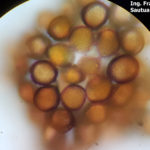
- Uredosporas de Puccinia sorghi. Autor: Dr. Francisco Sautua
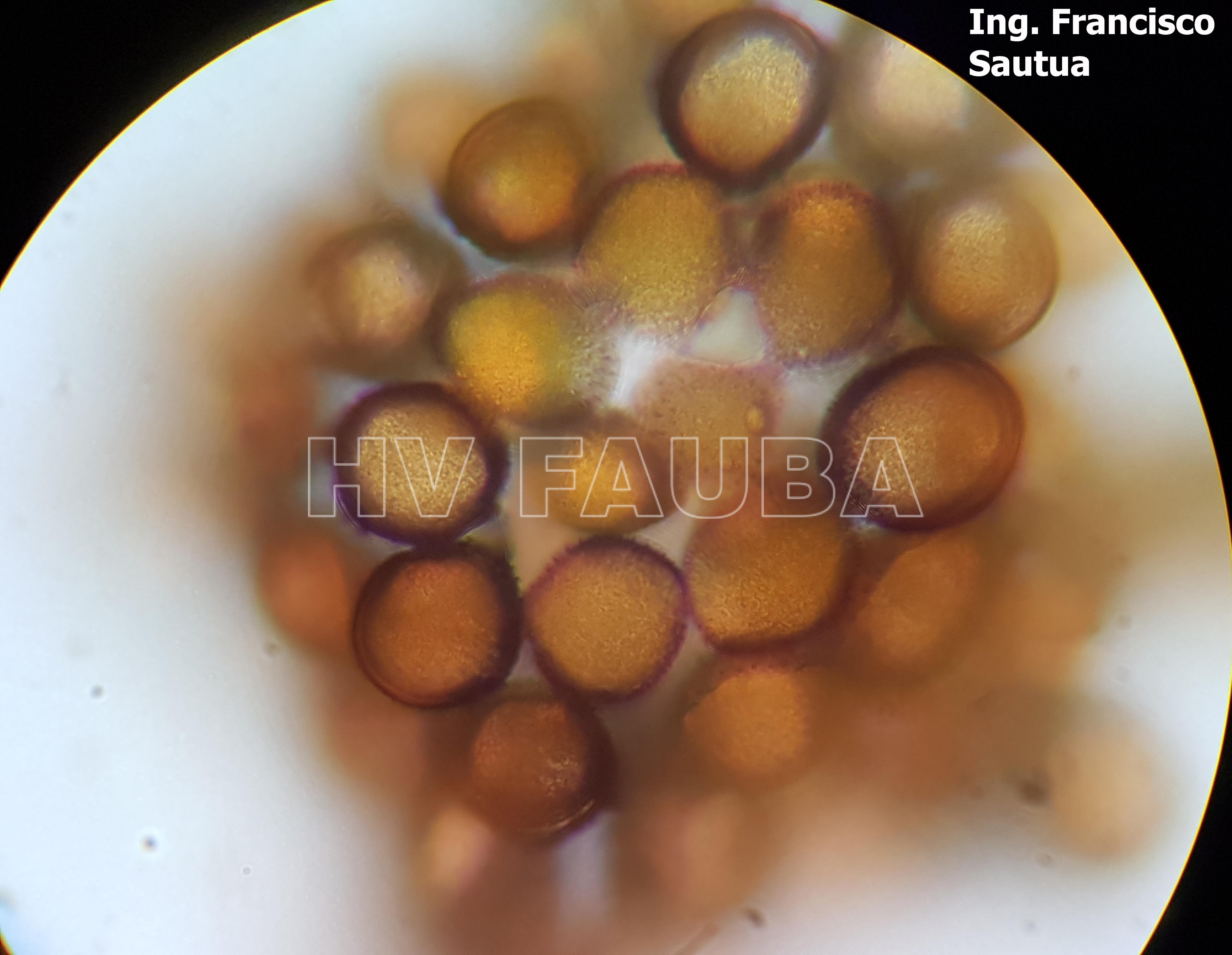
-
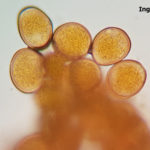
- Uredosporas de Puccinia sorghi. Autor: Dr. Francisco Sautua

-

- Signo (pústulas) de la Roya común del maíz. Autores: Dra. Verónica Felipe, Ing. Francisco Sautua. Villa María, Córdoba, 2020.

-

- Síntomas y Signo (pústulas) de la Roya común del maíz. Autores: Dra. Verónica Felipe, Ing. Francisco Sautua. Villa María, Córdoba, 2020.

-

- Autor: Laura Galvez

-

- Autor: Laura Galvez

-

- Autor: Laura Galvez

-

- Autor: Laura Galvez

.
-

- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua

-
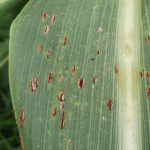
- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua
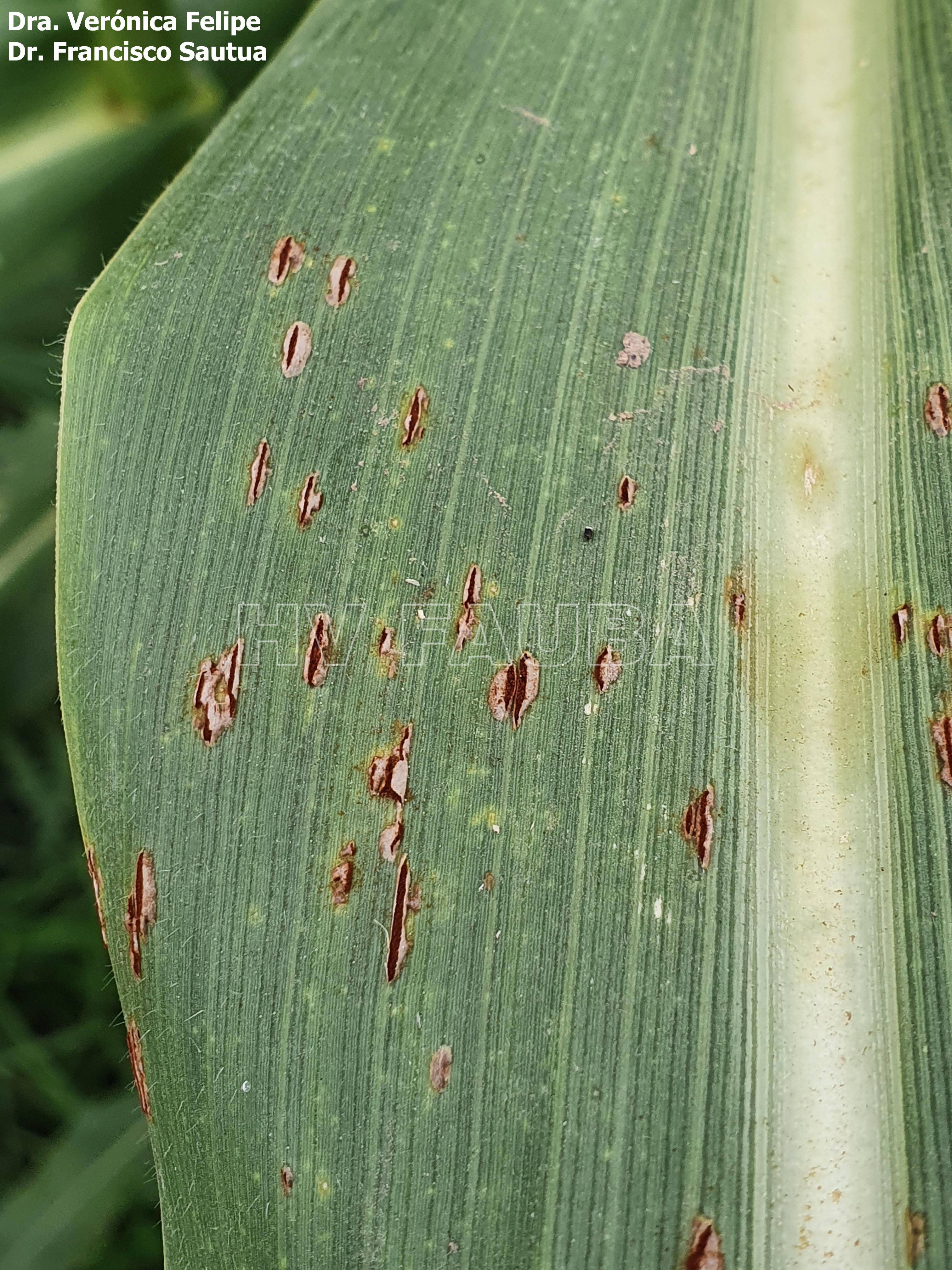
-

- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua

-

- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua

-

- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua

-

- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua

-

- Autores: Dra. Verónica Felipe, Dr. Francisco Sautua

.
.
Videos
Rust: the fungi that attacks plants. Created by Chris Hammang, Producer Sean O’Donoghue, Scientific Consultant Peter Dodds. C SIRO (Video)
.
Bibliografía
Antonelli (2012) La Roya Común del Maíz en Argentina (1942-2011). Memoria Técnica de la Chacra Experimental Miramar, Cultivos de Verano 2011/12.
Bade CIA, Carmona MA (2011) Comparison of methods to assess severity of common rust caused by Puccinia sorghi in maize. Tropical Plant Pathology 36(4): 264-266. doi: 10.1590/S1982-56762011000400009
Balint-Kurti P (2019) The plant hypersensitive response: concepts, control and consequences. Molecular Plant Pathology 20: 1163-1178. doi: 10.1111/mpp.12821
Carmona MA, Sautua FJ (2021) ¿Cuándo es realmente necesario aplicar fungicidas en maíz? Agronomía & Ambiente Rev. Facultad de agronomía UBA. Link
Carmona M, Sautua F (2021) ¿Cuándo es realmente necesario aplicar fungicidas en maíz? 5º Congreso argentino de fitopatología, 59th Meeting of the APS Caribbean división. 22 y 23 de septiembre de 2021 (modalidad virtual).
Chavan S, Gray J, Smith SM (2015) Diversity and evolution of Rp1 rust resistance genes in four maize lines. Theoretical and Applied Genetics 128(5): 985-998. doi: 10.1007/s00122-015-2484-2
Collins N, Drake J, Ayliffe M, Sun Q, Ellis J, Hulbert S, Pryor T (1999) Molecular Characterization of the Maize Rp1-D Rust Resistance Haplotype and Its Mutants. Plant Cell 11(7): 1365–1376. doi: 10.1105/tpc.11.7.1365
Darino, Martín Alejandro (2015) Estudio de las resistencias a roya común del maíz y a roya de la hoja del trigo. Tesis Doctoral, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. 110 p. LINK
Darino MA, Rochi L, Lia VV, Kreff ED, Pergolesi MF, Ingala LR, Dieguez MJ, Sacco F (2016) Virulence Characterization and Identification of Maize Lines Resistant to Puccinia sorghi Schwein. Present in the Argentine Corn Belt Region. Plant Disease 100(4): 770-776. doi: 10.1094/PDIS-06-15-0639-RE
Dean R, Van Kan JAL, Pretorius ZA, Hammond-Kosack KE, Di Pietro A, Spanu PD, et al. (2012). The Top 10 fungal pathogens in molecular plant pathology. Molecular Plant Pathology 13: 414–430. doi: 10.1111/j.1364-3703.2011.00783.x
Godoy EF, Bruni O (1952) Ciclo evolutivo de las royas del lino (Melampsora lini) y del maíz (Puccinia sorghi) en la Argentina. Revista Argentina de Agronomía 19: 21-34.
Gonzalez MP, Eyherabide G, Laguna IG (2011) Variabilidad de Puccinia sorghi en la zona maicera núcleo Argentina. Tropical Plant Pathology 36(3): 195-199. doi: 10.1590/S1982-56762011000300009
Guerra FA, Brücher E, De Rossi RL, Plazas MC, Guerra GD, Ducasse DA (2016) First Report of Oxalis conorrhiza as Alternate Host of Puccinia sorghi, Causal Agent of Common Rust of Maize. Plant Disease 100(2): 519. doi: 10.1094/PDIS-05-15-0506-PDN
Guerra FA, De Rossi RL, Brücher E, et al. (2018) Occurrence of the complete cycle of Puccinia sorghi Schw. in Argentina and implications on the common corn rust epidemiology. European Journal of Plant Pathology 1–7. doi: 10.1007/s10658-018-01645-3
, , , The maize ZmMIEL1 E3 ligase and ZmMYB83 transcription factor proteins interact and regulate the hypersensitive defence response. Molecular Plant Pathology 00: 1– 16. doi: 10.1111/mpp.13057
, , , et al. (2021) Analysis of the transcriptomic, metabolomic, and gene regulatory responses to Puccinia sorghi in maize. Molecular Plant Pathology 22: 465– 479. doi: 10.1111/mpp.13040
Lintz J, Dubrulle G, Cawston E, et al. (2022) A Short Review of Anti-Rust Fungi Peptides: Diversity and Bioassays. Front. Agron. 4: 966211. doi: 10.3389/fagro.2022.966211
MunkvoldGary P, WhiteDonald G (2015) Compendium of Corn Diseases, Fourth Edition. Diseases and Pests Compendium Series. American Phytopathological Society Press, St. Paul, MN. doi: 10.1094/9780890544945
Pataky JK, Pate MC, Hulbert SH (2001) Resistance genes in the rp1 region of maize effective against Puccinia sorghi virulent on the Rp1-D gene in North America. Plant Disease 85: 165-168. doi: 10.1094/PDIS.2001.85.2.165
Pryor T (1994) Maize and Puccinia sorghi: A System for the Study of the Genetic and Molecular Basis of Host-Pathogen Interactions. In: Freeling M, Walbot V (eds) The Maize Handbook. Springer Lab Manuals. Springer, New York, NY. doi: 10.1007/978-1-4612-2694-9_36
Rochi L, Diéguez MJ, Burguener G, Darino MA, Pergolesi MF, Ingala LR, Cuyeu AR, Turjanski A, Kreff ED, Sacco F (2016) Characterization and comparative analysis of the genome of Puccinia sorghi Schwein, the causal agent of maize common rust. Fungal Genetics and Biology (In Press, Corrected Proof). doi: 10.1016/j.fgb.2016.10.001
Sucher J, Boni R, Yang P, Rogowsky P, Büchner H, Kastner C, Kumlehn J, Krattinger SG, Keller B (2017) The durable wheat disease resistance gene Lr34 confers common rust and northern corn leaf blight resistance in maize. Plant Biotechnology Journal 15(4): 489-496. doi: 10.1111/pbi.12647
Webb CA, Richter TE, Collins NC, Nicolas M, Trick HN, Pryor T, Hulbert SH (2002) Genetic and Molecular Characterization of the Maize rp3 Rust Resistance Locus. Genetics 162(1): 381–394. Online ISSN 1943-2631
Yakovleva N, Ramirez-Cabral Z, Kumar L, Shabani L (2017) Global risk levels for corn rusts (Puccinia sorghi and Puccinia polysora) under climate change projections. Journal of Phytopathology 165(9): 563–574. doi: 10.1111/jph.12593