.
Condición fitosanitaria: ?
Grupo de cultivos: Hortícolas
Especie hospedante:
papa (Solanum tuberosum L.)
tomate (Lycopersicon esculentum Dunal L.)
.
Rango de Hospedantes: Solanáceas. Solanum demissum, S. caripense, S. nigrum-americanum, S. chacoense, S. colombianum, S. microdontum, S. platypterum, S. spegazzinii (Reddick, 1932; Buritica et al., 1968)
Kern (1933) y Pardo-Cardona (2002) describen e ilustran hongos de roya adicionales reportados en especies de Solanum, además de papa cultivada. Estos se diferencian principalmente por diferencias menores en la morfología de las teliosporas. Puccinia solani-tristis se reportó en varias especies silvestres de Solanum en Brasil (Mendes et al., 1998); este patógeno y otras ocho especies de Puccinia se reportaron en Colombia (Pardo-Cardona, 2002).
Epidemiología: policíclica, subaguda.
Ciclo: microcíclica, autoica. No se conoce ningún hospedante intermediario.
Etiología: Hongo. Biotrófico
Agente causal: Puccinia pittieriana Henn., 1904
Taxonomía: Fungi > Basidiomycota > Pucciniomycotina > Pucciniomycetes > Pucciniales > Pucciniaceae > Puccinia > P. pittieriana
.
La roya común es la única roya que afecta al tomate; pero en papa hay dos royas, la roya común y la roya deformante (causada por Aecidium cantensis).
Es una roya microcíclica, autoica. Produce sólo teliosporas que normalmente germinan in situ y basidiosporas que re-infectan las hojas de papa o tomate.
No existe información sobre formae speciales y razas.
.
-
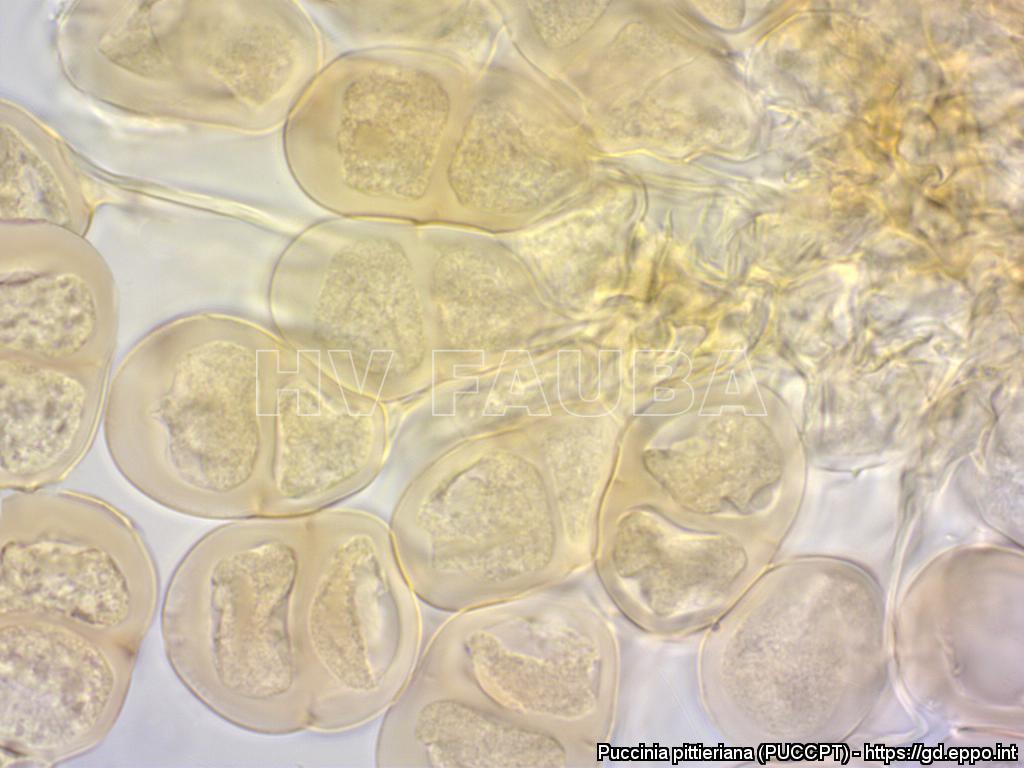
- Teliosporas de P. pittieriana. Autor: Mónica A. G. Otálora, Institute of Integrative Biology, ETH Zurich (CH), publicado en EPPO Database
-
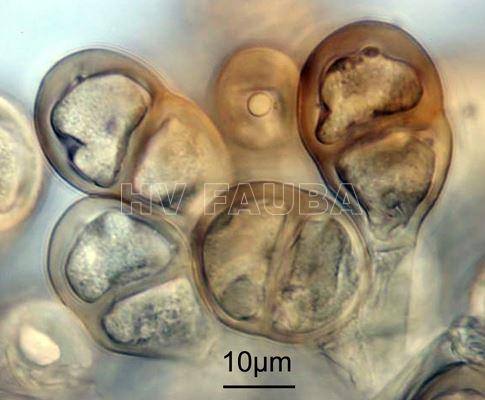
- Teliosporas de Puccinia pittieriana (x1000). Autor: USDA-ARS/Systematic Mycology & Microbiology Laboratory
.
.
Importancia económica
Las pérdidas de rendimiento/calidad debidas a la roya común de la papa no han sido cuantificadas en ninguno de los países en los que se ha informado sobre la enfermedad. P. pittieriana solo parece ser un factor limitante para la producción de papa en el norte de Ecuador, a veces en Colombia y rara vez en Perú (CABI, 2015). Las mayores pérdidas se reportaron en el norte de Ecuador cerca de la línea ecuatorial, donde se produce papa en un área de meseta en las provincias de Carchi y Tungurahua. Partes considerables de esta zona se encuentran por encima de los 3.000 m de altitud, con condiciones muy favorables para el desarrollo de la enfermedad (Velastegui, 1991). Ocasionalmente se reportaron pérdidas serias en Colombia en los Departamentos de Nariño, Caldas y Tolima (Chardon y Toro, 1930; Castaño, 1952). Sin embargo, como la literatura que se refiere a los impactos del patógeno es muy antigua, existe incertidumbre sobre las pérdidas de rendimiento/calidad que actualmente provoca el patógeno en los países infestados.
.
Distribución geográfica
Puccinia pittieriana es autóctona de América Central y del Sur, donde se encuentra en valles montañosos restringidos de las tierras altas frías (Bolivia*, Brazil, Colombia, Costa Rica, Ecuador, Mexico, Peru, Venezuela). La enfermedad es más común en altitudes de 3000 a 4300 m, aunque también se ha informado en un valle más bajo y cálido de Perú, a una altura de 2700 m (French, 1981). P. pittieriana no ha sido reportada en otros continentes.
* el reporte de Bolivia en Solanum platypterum es considerado dudoso. Se considera que existe incertidumbre sobre la presencia del patógeno en Paraguay y Bolivia.
.
-
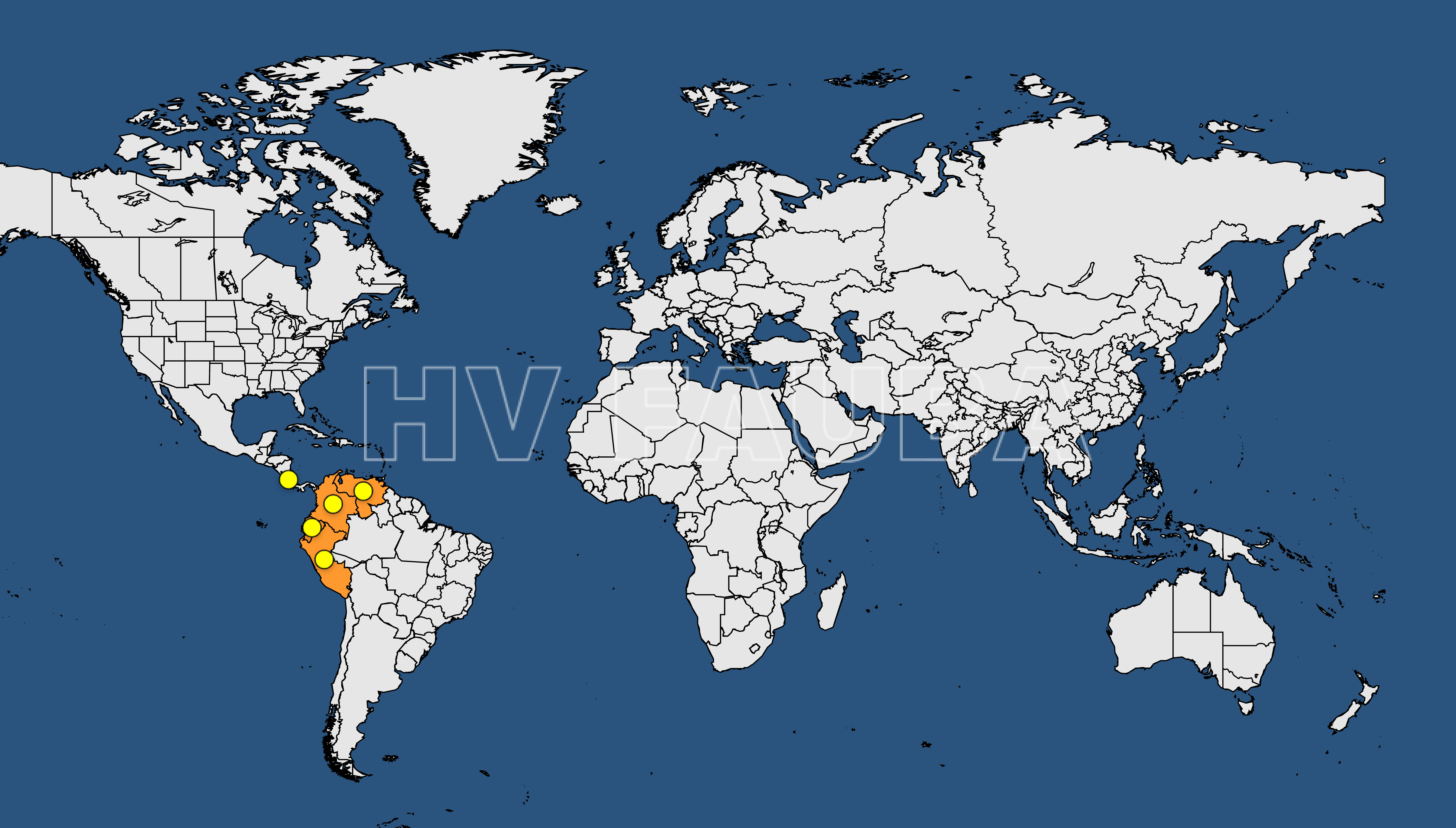
- Distribución mundial de la roya de la papa y el tomate, causada por Puccinia pittieriana. Fuente: https://gd.eppo.int/taxon/PUCCPT/distribution
.
.
Sintomatología
Con excepción de las semillas, todas las partes aéreas de las plantas de papa y tomate (tallos, hojas, pecíolos, flores y frutos) pueden verse afectadas por el patógeno (French, 2001). No se sabe que el patógeno se presente en tubérculos, semillas verdaderas, plántulas o plantas hospederas micropropagadas (CABI, 2015). La enfermedad se caracteriza porque en el haz de los foliolos, se presentan hendiduras más o menos cloróticas y en el envés, aparecen pústulas de color marrón rojizo de aproximadamente 3 a 5 mm de diámetro. Estas pústulas sobresalen 1-3 mm, con las correspondientes depresiones en la superficie superior de la hoja, y pueden estar rodeadas de halos cloróticos o necróticos. Los síntomas aparecen primero en el envés de las hojas como manchas diminutas, redondas (ocasionalmente alargadas), de color blanco verdoso, de 3 a 4 mm de diámetro, aunque su tamaño varía según la especie hospedante de Solanum y posiblemente la raza del patógeno (Castaño, 1952). Algunas lesiones se alargan y sus ejes más largos alcanzan los 8 mm (French, 1981, 2001). Luego se vuelven de color crema, con centros rojizos, luego rojo tomate y, finalmente, rojo óxido a marrón café. Posteriormente, las lesiones sobresalen de 1 a 3 mm, con las correspondientes depresiones en la superficie superior de la hoja, y pueden estar rodeadas de halos cloróticos o necróticos. La defoliación se produce cuando se forman cientos de lesiones en una hoja. Se producen lesiones alargadas o irregulares en pecíolos y tallos (French, 2001; Kern, 1933; Laundon y Rainbow, 1971).
No hay información sobre frutos de tomate afectados por el patógeno.
En papa, las dos royas provocan diferentes síntomas y signos (fructificaciones): la roya común de la papa produce telia típicos en lesiones de hojas y tallos, mientras que la roya deformante produce aecia en forma de platillo y causa distorsión de hojas y tallos (French, 2001).
.
-

- Pústulas sobresalientes de P. pittieriana en el envés de hoja de papa. Fuente: www.cabidigitallibrary.org
.
Ciclo de la enfermedad
El patógeno sobrevive como teliosporas en cultivos de papa superpuestos, en malezas solanáceas y/o en plantas voluntarias. Las teliosporas pueden persistir en los restos de plantas y en el suelo adherido a los tubérculos de papa, pero no se ha determinado la longevidad de su supervivencia (EPPO, 1988). In vitro y a temperaturas inferiores a 15°C, las teliosporas germinan en 1 h para producir un basidio (promicelio), que da lugar a cuatro basidiosporas (esporidios) dentro de 3 a 24 h. A temperaturas superiores a 15°C, el basidio suele seguir creciendo vegetativamente sin formar basidiosporas (CABI, 2015). Cuando se desprenden, las basidiosporas germinan inmediatamente para infectar plantas hospedantes susceptibles. A temperaturas ≤ 16°C, los primeros síntomas (lesiones) aparecen en 14–16 días en plantas de papa. Las lesiones se desarrollan por completo dentro de 20 a 25 días. Las teliosporas maduran entre 30 y 40 días después de la inoculación (French, 1981, 2001). Temperaturas promedio alrededor de 10°C con 10–12 h de mojado (humedad libre sobre la superficie de las plantas) son necesarias para el desarrollo de la enfermedad y la propagación del patógeno (French, 1981, 2001; CABI, 2015). El inóculo (basidiosporas) producido en cultivos hospedantes sembrados anteriormente o plantas hospedantes silvestres, es diseminado por el viento (Laundon y Rainbow, 1971).
P. pittieriana puede propagarse mediante basidiosporas diseminadas por el viento. Sin embargo, el potencial de P. pittieriana para propagarse por el viento es menor que el de otros hongos causantes de roya que producen urediniosporas (ausentes en P. pittieriana), debido a la limitación para la dispersión a larga distancia de las basidiosporas (French, 2001). Las basidiosporas tienen una vida corta y no se producen en grandes cantidades ni a temperaturas superiores a 15°C. El patógeno también podría propagarse por el movimiento de material vegetal vivo o muerto infectado (p. ej., residuos de cultivos), o con los residuos de cultivos o el suelo infestado por teliosporas y acompañando el movimiento/comercio de tubérculos de papa (EPPO, 1988).
.
.
Medidas de manejo
En países en los que la enfermedad no está presente:
Medidas para prevenir la entrada del patógeno en un área:
* Obtención material vegetal de áreas libres del patógeno;
* Certificado fitosanitario para la exportación de material vegetal de países infestados;
* Inspección del material vegetal antes de la exportación y en el punto de entrada del país importador.
.
En países en los que la enfermedad está presente (
Medidas para prevenir el establecimiento y la propagación del patógeno en un área:
* Restringir el movimiento de material vegetal infectado, incluidos los residuos de cultivos y el suelo adherido a los tubérculos de papa.
* Se encontró resistencia genética a la roya común cuando se examinaron 136 cultivares de papa mediante infección natural en el campo en dos lugares de la provincia de Tungurahua, Ecuador. Se reportó que un total de 12 cultivares tenían resistencia. Las tres más resistentes fueron la Colección Ecuatoriana de Papa números 305, 314 y 503 (Coronel-Orijalva, 1970).
* Control químico (aplicación de fungicidas)
* Manejo de residuos de cultivos
.
.
Bibliografía
Alandia-Borda S (1966) Nuevas identificaciones de hongos parasiticos de plantas economicas de Bolivia. Turrialba 16: 398–401.
, (1968) La roya de la papa en Colombia y sus implicaciones. Agricultura Tropical (Colombia) 24: 221–222.
CABI (2015) Invasive species compendium. Link
(1952) Roya de la papa. Agricultura Tropical (Bogota) 8: 47–48.
(1930) Mycological explorations of Colombia. Journal of Department of Agriculture Puerto Rico 14: 195–369.
Coronel-Orijalva MA (1970) Determination of resistances to late blight and common rust and other characteristics of potato germplasm). Tesis de grado (Ing. Agr.) Universidad Central del Ecuador (Quito). 80 pp.
EFSA Panel on Plant Health (PLH), et al. (2017) Pest categorisation of Puccinia pittieriana. EFSA J. 15(11): e05036. doi: 10.2903/j.efsa.2017.5036
EPPO (European and Mediterranean Plant Protection Organization) (1988) Datasheets on quarantine organisms 155, Puccinia pittieriana. Bulletin OEPP/EPPO Bulletin 18: 517–519.
French ER (1981) Common rust. In: Compendium of Potato Diseases. APS Press, Saint Paul, Minnesota, USA, 65 pp.
French ER (2001) Common rust. In: Compendium of Potato Diseases (2nd Edition). Minnesota, USA, APS Press, Saint Paul. pp. 20–21.
(2001) Deforming rust. In: Compendium of Potato Diseases. 2nd Edition. APS Press, Saint Paul, Minnesota, USA, 21 pp.
(1933) The microcyclic species of Puccinia on Solanum. Mycologia 25: 435–441.
Laundon GF, Rainbow AF (1971) Descriptions of Fungi and Bacteria: Puccinia pittieriana. IMI Descriptions of Fungi and Bacteria 29, Wallingford, UK, CAB International.
, , , (1998) Fungos em plantas no Brasil. Embrapa-SPI/Embrapa-Cenargen, Brasilia, DF, Brazil. 555 pp.
(2002) Especies microciclicas de Puccinia Persoon sobre Solanum L. in Colombia. Caldasia 23: 427–434.
(1932) Some diseases of wild potatoes in Mexico. Phytopathology 22: 609–612.
(1991) Control quimico de Puccinia pittieriana P. Henn., agente de la roya de la papa, con fungicidas sistemicos. In Memorias del V seminario Nacional de Sanidad Vegetal, 7–9 Junio 1988, Universidadde Cuenca, Ecuador, pp. 104–105.