.
Condición fitosanitaria: Plaga Cuarentenaria Ausente
Grupo de cultivos: Frutales de pepita
Especie hospedante: Manzano (Malus domestica)
Rango de hospedantes: relativamente estrecho
Epidemiología: es considerada una enfermedad poliética
Etiología: Bacteria sin pared celular (Clase Mollicutes). Limitada al floema. Considerada Biotrófica. Bacteria fastidiosa (no cultivable).
Agente causal: Candidatus Phytoplasma mali Seemuller and Schneider 2004, candidatus name (nombre provisional para organismos bien caracterizados pero aún sin cultivar in vitro)
Taxonomía: Bacteria > Terrabacteriagroup > Tenericutes > Mollicutes > Acholeplasmatales > Acholeplasmataceae > Candidatus Phytoplasma > 16SrX (Apple proliferation group)
.
Los Candidatus Phytoplasma son procariotes sin pared celular pertenecientes a la clase Mollicutes (mycoplasmas), que invaden el floema y se transmiten por chicharritas vectoras. Los fitoplasmas no pueden ser cultivados in vitro, y se caracterizan por poseer un genoma pequeño (0.53 – 1.35 Mb), con bajo %GC (21-28%) (Kube et al. 2008; Marcone et al. 1999). Causan amarillamientos, achaparramientos, escobas de bruja y otras alteraciones del crecimiento.
El agente causal de la Proliferación del manzano se encuentra en los tubos cribosos del floema. El fitoplasma es altamente pleomórfico, de aproximadamente 200-800 nm de diámetro, limitado por una membrana citoplásmica trilaminar pero sin una pared celular rígida (Seemüller, 1990).
.
.
Antecedentes
Se ha registrado en Austria, Bélgica, Bulgaria, Grecia, Noruega, Rumania, Suiza, antigua URSS, antigua Yugoslavia (Németh, 1986), India y Sudáfrica (Seemüller, 1990), pero en base a síntomas, por lo que se requiere confirmación adicional. La Proliferación del manzano es considerada una de las enfermedades causadas por fitoplasmas más importantes del manzano, particularmente en las áreas norteñas de la región sur de Europa.
.
Síntomas
Las hojas de las plantas infectadas ruedan hacia abajo y se vuelven quebradizas, están dentadas fina e irregularmente y son más pequeñas de lo normal. En otoño las hojas se vuelven rojas en contraste con la coloración amarilla de las plantas sanas. En verano las hojas suelen ser cloróticas. La defoliación temprana puede ocurrir. Una roseta de hojas terminales a veces se desarrolla al final de la temporada en lugar de brotes latentes normales. Las estípulas se agrandan anormalmente, mientras que los pecíolos son bastante cortos.
Los brotes se desarrollan prematuramente a partir de brotes axilares y dan lugar a brotes secundarios que forman una escoba de bruja. El ángulo entre los brotes secundarios y el brote principal es anormalmente estrecho. La roseta de hojas puede aparecer en los extremos de los brotes o las puntas de los brotes pueden morir hacia atrás.
En algunos casos, las flores muestran numerosos pétalos y los pedúnculos son anormalmente largos. Pueden permanecer en el árbol durante un largo período. Las frutas se reducen en tamaño, y poseen coloración incompleta y mal sabor. Los síntomas se distribuyen de manera desigual en toda la planta, a menudo se encuentran ramas sanas con frutos normales. Los árboles afectados son menos vigorosos, pero rara vez mueren. A veces, después de una fase de choque, los árboles pueden producir normalmente, especialmente si se fertilizan adecuadamente. El sistema de raíces fibrosas de los árboles infectados forma masas compactas similares a un fieltro de raíces cortas, de modo que los más grandes no pueden desarrollarse. El peso de la raíz se reduce en un 20-40%. La circunferencia del tronco y el diámetro de la corona se reducen en comparación con los árboles sanos (Kunze, 1979).
.
-

- 01 Manzano cv. Golden Delicious con desarrollo prematuro de brotes axilares que dan como resultado una escoba de bruja cerca del vértice del brote principal en primavera, causado por Candidatus Ca Phytoplasma mali. Autor: EPPO Global Database

-

- 02 Hojas de árboles infectados con Candidatus Ca Phytoplasma mali (izquierda), son más pequeñas que las normales (derecha) y tienen grandes estípulas en la base del tallo. Autor: EPPO Global Database

-

- 03 Hoja de manzano cv. Golden Delicious, mostrando tamaño reducido de la lámina y estípulas agrandadas; Hoja sana a la izquierda. Autor: EPPO Global Database

-

- 04 Síntomas en frutos de manzano infectados con Candidatus Ca Phytoplasma mali (derecha), son más pequeños que las normales (izquierda). Autor: EPPO Global Database

-

-
05 Síntomas de la Proliferación del manzano en Malus x domestica cv. Golden Delicious, causados por diferentes cepas del fitoplasma: [(A) acceso 1/93 avirulento; (B) acceso 12/93 virulento] y plantas de Catharanthus roseus [(C) sanas; (D) acceso 1/93 avirulento; (E) Adhesión 12/93 virulenta]. Autor: Zimmermann et al., 2015.
-
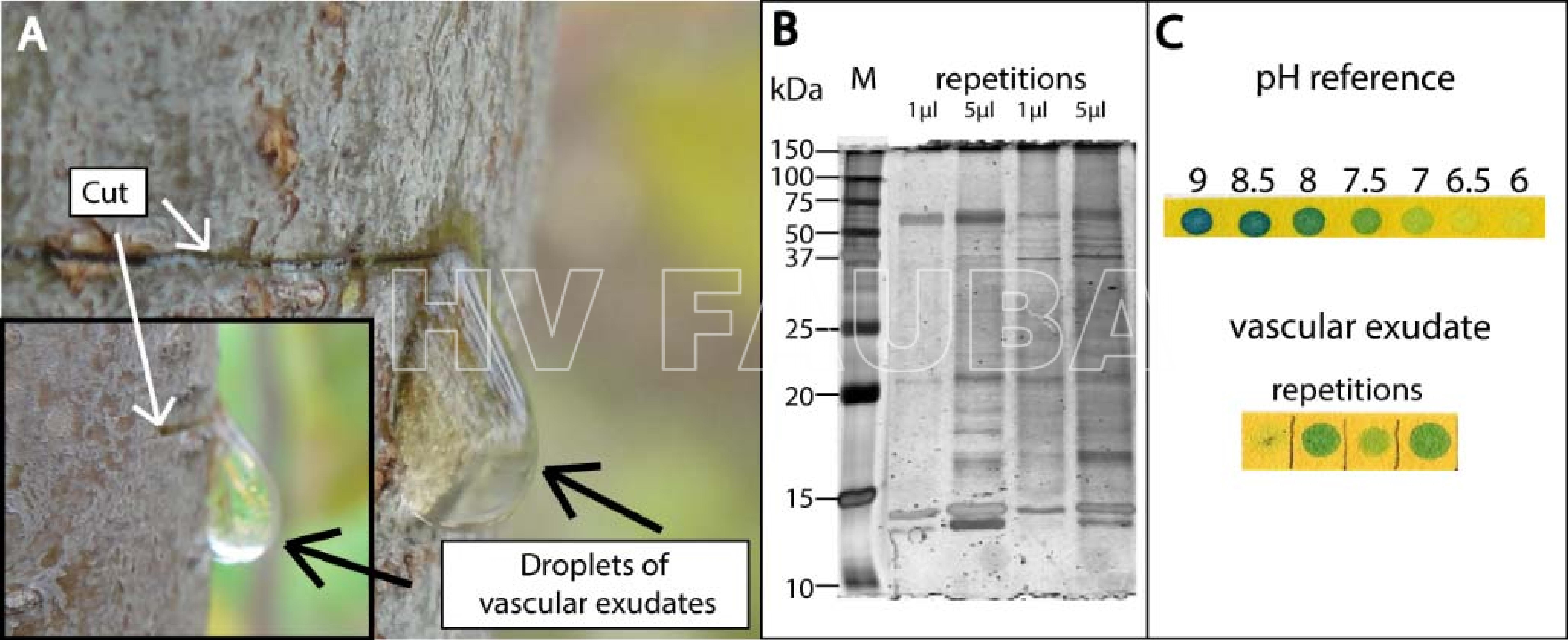
- Manzano infectado con Candidatu P. mali mostrando exudación vascular inducida por la herida. El gran volumen (~300 µl) por gota (A), las numerosas proteínas (B) y el pH citoplásmico de 7.5‐8 (C) indican el contenido del tubo criboso. El exudado se recogió del tallo de un manzano después de cortarlo con una pipeta y se transfirió a un tampón de muestra reductor concentrado 4 veces (Roti-Load 1; Carl-Roth, Karlsruhe, Alemania) en la proporción de 1: 3. Se realizó un SDS-PAGE unidimensional de exudado de acuerdo con Laemmli (1970) utilizando un gel de apilamiento al 4% y un gel de separación al 12% en un sistema de electroforesis MiniProtean 3 (Bio-Rad Laboratories, Hercules, CA, EE. UU.) Con precisión Plus Protein Standard – All Blue (Bio-Rad) como marcador de tamaño de proteína. Los geles se tiñeron con plata (Heukeshoven y Dernick 1988). Los exudados (V = 1–4 µL) se colocaron en papel indicador (Macherey-Nagel, Düren, Alemania). Se utilizaron tampones MES / TRIS con diferentes valores de pH (4-9.7) para la línea de calibración. Autor: Zimmermann et al., 2015
.
Diagnóstico
Ensayo biologico
La identificación positiva requiere la transmisión a una especie leñosa indicadora. Malus dawsoniana, una planta indicadora muy sensible, cuando se injerta en verano en el vástago, desarrolla síntomas el otoño siguiente. Usando la técnica de doble brote, la reacción aparece después de la brotación (Anon., 1997). El uso del reactivo DAPI (1,6 diamidino 2-fenilindol) puede ayudar a detectar la fluorescencia de los fitoplasmas en los tubos cribosos del floema bajo la corteza de los manzanos infectados
Ensayo serológico
La Proliferación del manzano (PM) puede detectarse en manzanos infectados utilizando anticuerpos monoclonales (MAbs) para fitoplasmas PM obtenidos de Catharanthus roseus infectados con fitoplasma PM como fuente de antígeno. Estos MAbs reaccionan específicamente en ELISA e inmunofluorescencia (IF) (Loi et al., 1998). Su patente está pendiente. ADGEN Ltd. también ha producido un antisuero específico para el fitoplasma PM, que funciona con extractos de manzana si hay suficiente antígeno presente. Normalmente la concentración de fitoplasma es un factor limitante.
Ensayo Molecular
Se han desarrollado varios primers para el diagnóstico de enfermedades causadas por fitoplasmas, entre ellos los específicos del grupo de la Proliferación del manzano (PM) (Lee et al., 1993, 1995; Firrao et al., 1994; Jarausch et al., 1994 Lorenz et al., 1995; Smart et al., 1996; Brzin et al., 2003; Torres et al., 2005; Baric et al., 2011).
.
-
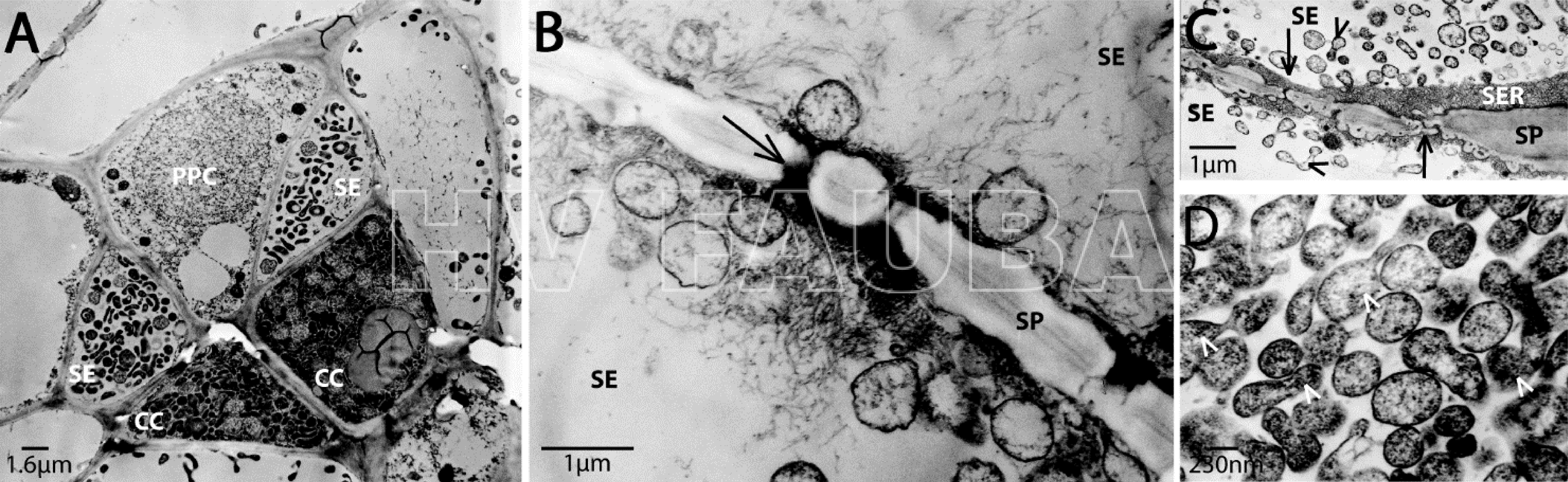
- Micrografías electrónicas de transmisión de Candidatus P. mali infectando plantas de Cuscuta odorata. La descripción general de las células del floema (A) muestra una infección de tubos cribosos (SE), células compañeras (CC) y células del parénquima del floema (PPC) con fitoplasmas. Un mayor aumento o zoom (B, C) ilustra la acumulación de fitoplasmas en los tubos cribosos y los fitoplasmas densamente empaquetados, algunos son brotes (puntas de flecha) (D). Los poros del tamiz (tubo criboso) están marcados con flechas. SP (sieve plate), placa de tamiz; SER (sieve element reticulum), retículo de elemento tamiz. Autor: Zimmermann et al., 2015
.
Patogenicidad y Factores de virulencia
A pesar de su importancia económica, poco se sabe sobre los mecanismos moleculares de la manifestación de la enfermedad dentro de los manzanos. Janik et al. (2017) identificaron dos factores de transcripción TCP (TEOSINTE BRANCHED / CYCLOIDEA / PROLIFERATING CELL FACTOR) de Malus x domestica como socios de unión del efector de tipo SAP11 ATP_00189 de Ca P. mali. En este estudio, los análisis de fitohormonas revelaron un efecto de la infección por Ca P. mali en los niveles de jasmonatos, ácido salicílico y ácido abscísico, mostrando que Ca P. mali afecta los niveles fitohormonales en los manzanos, lo que está en línea con las funciones del efector asumido de su unión al factor de transcripción TCP. Según los autores, esta es la primera caracterización de los objetivos moleculares de un efector de Ca P. mali y, por lo tanto, proporciona la base para comprender mejor el desarrollo de los síntomas y el progreso de la enfermedad durante la Proliferación del manzano. Como los homólogos de SAP11 se encuentran en varias especies de fitoplasmas que infectan una amplia gama de plantas diferentes, las proteínas similares a SAP11 parecen ser actores clave en la infección del fitoplasma.
Si bien en la actualidad el estudio de Janik et al. (2017) representa la única prueba molecular o bioquímica directa que identifique las proteínas de virulencia o los efectores de Candidatus Phytoplasma mali, existe evidencia circunstancial de que las proteínas de membrana de Ca. P. mali tienen una influencia significativa en la patogenicidad. Estos hallazgos se basan en el monitoreo de los datos de la enfermedad en ensayos a largo plazo de accesiones PM de diferente origen y datos moleculares de cepas de patógenos asociados derivadas. Sobre la base de los síntomas fenotípicos, las accesiones se clasificaron como fuertemente, moderadamente o levemente virulentas (Seemüller y Schneider 2007; Seemüller et al. 2011). Los datos moleculares provienen de un grupo de genes, que codifican proteínas AAA+ que están presentes en un número inusualmente alto en fitoplasmas, como por ejemplo los genes de ATPasas hflB y AAA+. El análisis de las secuencias de ácidos nucleicos o secuencias de aminoácidos derivadas de algunos de los genes reveló sustituciones relacionadas con la virulencia. El análisis de conglomerados separó claramente las cepas virulentas de las no virulentas. La superfamilia AAA+ es un grupo grande, funcionalmente diverso de proteínas y comprende varios tipos de ATPasas (por ejemplo, ClpC, ClpV, p97) y proteasas (por ejemplo HflB, Grupo de Clp, Lon) que poseen el módulo ATPase (Langklotz et al. 2012; Snider et al. al. 2008). Ambas son proteínas integrales de membrana con el largo dominio C-terminal relevante catalítico que enfrenta el citosol. Sin embargo, la predicción de la topología indicó que la cola terminal C de cuatro ATPasas AAA+ y dos de los tres HflBs de Ca. P. mali se orientaban hacia el exterior, es decir hacia el citoplasma del tubo criboso (Seemüller et al. 2013). Las proteínas AAA+ son de una importancia crucial para la virulencia bacteriana. La ATPasa ClpC de Staphylococcus aureus, por ejemplo, regula la transcripción, que es un factor de virulencia importante (Luong et al. 2011). Del mismo modo, las proteínas AAA+ son esenciales para la función de los sistemas de secreción. Varias especies bacterianas como Salmonella enterica, Agrobacterium tumefaciens y Pseudomonas syringae utilizan el sistema de secreción tipo tres (TTSS) para inyectar factores de virulencia en el hospedante eucariótico (Van Melderen y Aertsen 2009; Alix y Blanc‐ Potard 2008). Este hallazgo inusual en la orientación de la cola C terminal es un nuevo aspecto para comprender la patogenicidad del fitoplasma a nivel del tubo criboso. Es concebible que las poderosas proteínas AAA+ ataquen las estructuras o componentes de los elementos del tamiz o tubo criboso del floema, en particular si los patógenos están unidos a la membrana. La adherencia a las membranas del hospedante está bien establecida para la mayoría de los micoplasmas patógenos para humanos y animales, y se considera un requisito previo para la colonización e infección evidenciada por el daño causado a las membranas (Razin et al. 1998). Sin embargo, tal unión, que se basa en la interacción proteína / proteína, no está claramente establecida para los fitoplasmas.
.
-
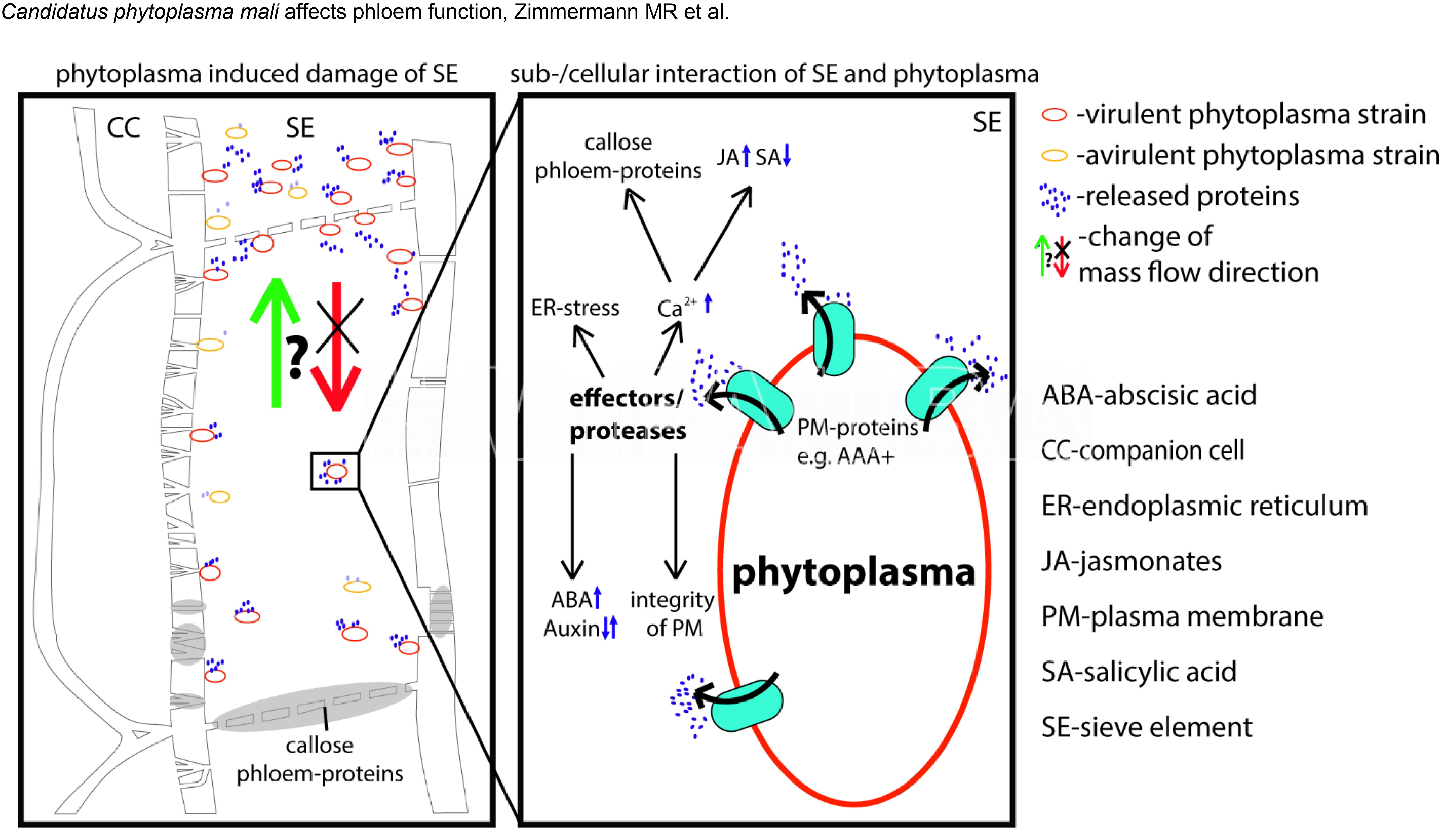
- Modelo propuesto por Zimmermann et al., 2015 sobre el nivel (sub)celular de interacción planta-fitoplasma.
.
Epidemiología
En el campo, la enfermedad parece propagarse naturalmente por los insectos vectores y por la fusión de raices. La Proliferación del manzano fue transmitida experimentalmente de manzano a manzano y a Catharanthus roseus por el vector Philaenus spumarius; de manzano a C. roseus por Aphrophora alni y Lepyronia coleoptrata; y de plantas de apio infectadas a plántulas de manzano por Arhianus interstitialis (Hegab y El-Zohairy, 1986). Estos experimentos demostraron que las ninfas pueden adquirir el patógeno y transmitirlo en la etapa adulta. Se encontró que los adultos mantienen la capacidad de transmitir el fitoplasma hasta el final de su vida. El período de incubación en manzanos dura 1-2 años (Krczal et al., 1988; Krczal y Bliefernich, 1992).
También, el insecto vector Fiebierella florii fue considerado como un vector putativo del fitoplasma causante de la Proliferación del manzano. ADN de Ca Phytoplasma mali fue detectado en ADN total extraído de F. florii atrapado en montes con manzanos enfermos de proliferación (Vega et al., 1993; Bliefernich y Krczal, 1995). Además, recientemente se ha demostrado que los psílidos Cacopsylla costalis, C. mali y C. melanoneura son vectores (Grando et al., 1998; Jarausch et al., 2003; Tedeschi et al., 2003; Tedeschi y Alma, 2004 ) y, según Tedeschi y Alma (2004), los psílidos parecen ser los vectores más importantes para la Proliferación del manzano. Schmid (1975) informó que, en términos de propagación, el 73% de los árboles en un monte frutal de manzanos se infectaron durante un período de 12 años; observando un incremento anual promedio del 18%.
.
-

- Ejemplo de un vector de Candidatus Phytoplasma mali. Hembra adulta de Cacopsylla picta, nueva generación (emigrante) en Malus spp. Autor: Mayer y Gross, JKI.
.
La distribución del fitoplasma dentro del árbol infectado está restringida a los elementos funcionales del floema, con la colonización de las partes sobre el suelo siguiendo un patrón estacional. El fitoplasma desaparece en las partes del suelo durante el invierno, cuando se produce la inactivación completa de los tubos cribosos en las partes aéreas de los árboles frutales de pepita. Durante este período, el fitoplasma aún sobrevive en las raíces y en primavera comienza a recolonizar el tallo y se dispara después del desarrollo de un nuevo círculo del floema. El patógeno puede ser adquirido por insectos chupadores de savia, como los enumerados anteriormente, dentro de los cuales puede multiplicarse, circular a las glándulas salivales y ser expulsado durante las sondas de alimentación para infectar otras plantas. Una vez que se infecta un vector, conserva la capacidad de transmitir el fitoplasma durante toda su vida útil, y se ha demostrado que los vectores de psílidos adultos que aún pasan el invierno todavía llevan el fitoplasma la próxima primavera (Tedeschi et al., 2003; Tedeschi y Alma, 2004).
.
-
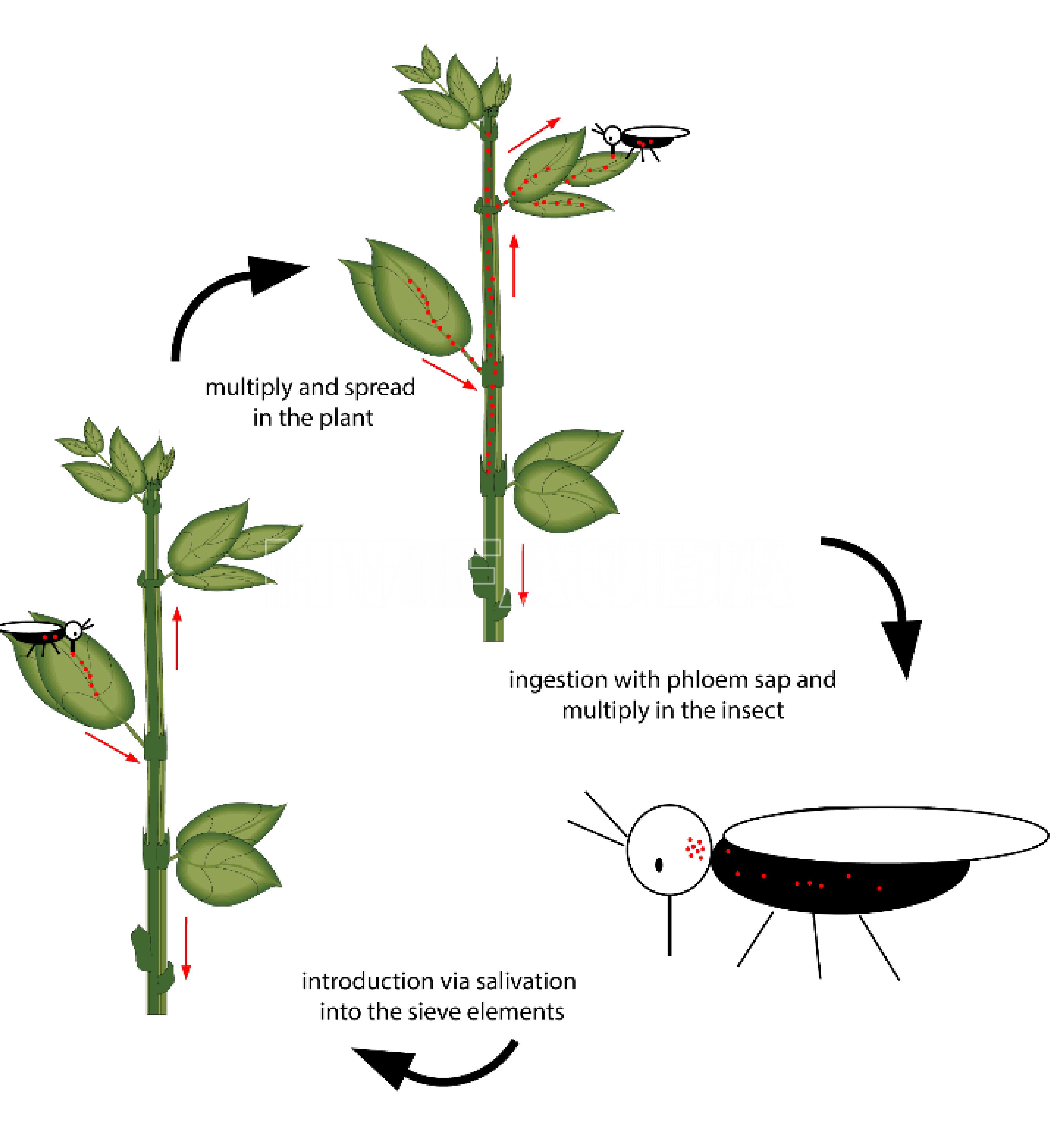
- Ciclo de vida del fitoplasma. Los puntos rojos representan los fitoplasmas y las flechas rojas muestran la dirección de propagación. Autor: Zimmermann et al., 2015
.
La temperatura parece tener un impacto significativo en la expresión de la enfermedad y, por lo tanto, en el impacto. Un estudio de Ducroquet et al. (1986) encontró que los síntomas de la Proliferación del manzano se desarrollaron a temperaturas de 21-24°C, pero no entre 29 y 32°C. La Proliferación del manzano es considerada una de las enfermedades causadas por fitoplasmas más importantes del manzano, particularmente en las áreas norteñas de la región sur de Europa, donde las temperaturas son las más propicias para la expresión de los síntomas. Fuera de esta región, donde ocurren condiciones de crecimiento más frías o más cálidas, la enfermedad parece tener menos importancia (Seemüller et al., 1998).
Desde el punto de vista epidemiológico, es considerada una enfermedad poliética.
.
Manejo preventivo de la enfermedad
Como medida preventiva, el uso de material de propagación sano es un requisito previo fundamental. El control intensivo de malezas y plagas de insectos en los montes frutales también es esencial. La eliminación de los retoños de raíz es una medida de control para reducir la difusión de los vectores que viven en las malezas. Los fitoplasmas son sensibles a la tetraciclina, pero este antibiótico es bacteriostático en lugar de bactericida. Después del tratamiento con antibióticos, los síntomas pueden disminuir notablemente durante 2 años (Casanova et al., 1980), pero a menudo esto solo se observa como un efecto temporal (Schmid, 1983). El uso de tetraciclina puede seleccionar involuntariamente para poblaciones bacterianas resistentes. En la actualidad, el uso de pie de injertos resistentes es la forma más eficiente de controlar la enfermedad (Seemüller, 1990).
.
.
.
Bibliografía
(2017), PM 7/62 (2) ‘Candidatus Phytoplasma mali’, ‘Ca. P. pyri’ and ‘Ca. P. prunorum’. EPPO Bull, 47: 146-163. doi: 10.1111/epp.12380
ProMED-mail. 2013. Apple proliferation – North America: First report, Canada. 0415.1646164. April 15, 2013. Last accessed May 2, 2013. Link
The IRPCM Phytoplasma/Spiroplasma Working Team – Phytoplasma taxonomy group. Int J Syst Evol Microbiol 54(4):1243-1255 doi: 10.1099/ijs.0.02854-0
Baric S, Dalla Via J (2004) A new approach to apple proliferation detection: a highly sensitive realtime PCR assay. Journal of Microbiological Methods 57: 135-145. doi: 10.1016/j.mimet.2003.12.009
Baric S, Kerschbamer C, Dalla Via J (2006) TaqMan real-time PCR versus four conventional PCR assays for detection of apple proliferation phytoplasma. Plant Molecular Biology Reporter 24: 169-184. doi: 10.1007/BF02914056
Baric S, Kerschbamer C, Dalla Via J (2007) Detection of latent apple proliferation infection in two differently aged apple orchards in South Tyrol (northern Italy). Bulletin of Insectology 60(2): 265-266. Link
Baric S, Kerschbamer C, Vigl J, Dalla Via J (2008) Translocation of apple proliferation phytoplasma via natural root grafts – a case study. European Journal of Plant Pathology 121: 207-211. doi: 10.1007/s10658-007-9256-z
Baric S, Berger J, Cainelli C, et al. (2011) Seasonal colonization of apple trees by ‘Candidatus Phytoplasma mali’ revealed by a new quantitative TaqMan real-time PCR approach. European Journal of Plant Pathology 129(3): 455-467. doi: 10.1007/s10658-010-9706-x
Baric S, Berger J, Cainelli C, et al. (2011) Molecular typing of ‘Candidatus Phytoplasma mali’ and epidemic history tracing by a combined T-RFLP/VNTR analysis approach. European Journal of Plant Pathology 131: 573. doi: 10.1007/s10658-011-9832-0
Ben Khalifa M, Fakhfakh H (2011) Detection of 16S rDNA of ‘Candidatus Phytoplasma mali’ in plum decline in Tunisia. Canadian Journal of Plant Pathology: 33(3): 332-336. doi: 10.1080/07060661.2011.599427
Bertaccini A, Paltrinieri S, Martini M, et al. (2008) Comparison of different detection systems for apple proliferation phytoplasmas in Trentino (North Italy). Acta Horticulturae 781: 453-458. doi: 10.17660/ActaHortic.2008.781.65
Bertaccini A, Weintraub PG, Rao GP, Mori N (2019) Phytoplasmas: Plant Pathogenic Bacteria – II. Springer. doi: 10.1007/978-981-13-2832-9
Bisognin C, Ciccotti AM, Moser M, et al. (2008) Establishment of an in vitro screening of apple proliferation-resistant rootstock genotypes based on micrografting. Acta Horticulturae 781: 375-380. doi: 10.17660/ActaHortic.2008.781.53
Bliefernich K, Krczal G (1995) Epidemiological studies on apple proliferation disease in Southern Germany. Acta Horticulturae 386: 444-453. Link
Brzin J, Ermacora P, Osler R, et al. (2003) Detection of apple proliferation by ELISA and PCR in growing and dormant apple trees. Journal of Plant Diseases and Protection 110: 476-483. doi: 10.1007/BF03356124
Canik D, Ertunc F (2007) Distribution and molecular characterization of apple proliferation phytoplasma in Turkey. Bulletin of Insectology 60(2): 335-336.
Carraro L, Ermacora P, Loi N, Osler R (2004) The recovery phenomenon in apple proliferation infected apple trees. Journal of Plant Pathology 86(2): 141-146. Link
Casanova R, Llacer G, Sanchez-Capuchino JA (1980) Remission of symptoms of apple proliferation, after injection of concentrated tetracycline solutions. Acta Phytopathologica Academiae Scientiarum Hungaricp, 15:273-277
Casati P, Quaglino F, Tedeschi R, et al. (2010) Identification and molecular characterization of ‘Candidatus Phytoplasma mali’ isolates in north-western Italy. Journal of Phytopathology 158: 81-87. doi: 10.1111/j.1439-0434.2009.01581.x
Ciccotti AM, Bianchedi PL, Bragagna P, et al. (2008) Natural and experimental transmission of ‘Candidatus Phytoplasma mali’ by root bridges. Acta Horticulturae 781: 459-464. doi: 10.17660/ActaHortic.2008.781.66
Cieslinska M, Morgas H (2011) Detection and identification of ‘Candidatus Phytoplasma prunorum’. ‘Candidatus Phytoplasma mali’, and ‘Candidatus Phytoplasma pyri’ in stone fruit trees in Poland. Journal of Phytopathol. 159: 217-222. doi: 10.1111/j.1439-0434.2010.01752.x
Davis R, Zhao Y, Dally EL, et al. (2013) ‘Candidatus Phytoplasma pruni’, a novel taxon associated with X-disease of stone fruits, Prunus spp.: multilocus characterization based on 16S rRNA, secY, and ribosomal protein genes. International Journal of Systematic and Evolutionary Microbiology 63: 766–776.
Ducroquet J-P, Dosba F, Lansac M, Mazy K (1986) Effet de la température sur l’expression des symptômes de la prolifération du pommier. Agronomie, 6(10):897-903.
Ermacora P, Carraro L, Martini M, et al. (2008) Apple proliferation susceptibility and sensitiveness in old apple-trees varieties naturally and artificially infected. Acta Horticulturae 781: 465-470. doi: 10.17660/ActaHortic.2008.781.67
Fernández F, et al. (2020) Enfermedades causadas por Fitoplasmas. Situación en Argentina. Boletín de la Asociación Argentina de Fitopatologos ISSN: 2618-1932. Diciembre 2020. N°7. Link
Fránová J, Ludvíková H, Paprštein F, Bertaccini A (2013) Genetic diversity of Czech ‘Candidatus Phytoplasma mali’ strains based on multilocus gene analyses. European Journal of Plant Pathology 136: 675-688. doi: 10.1007/s10658-013-0196-5
Fránová J, Lenz O, Přibylová J, et al. (2018) “Candidatus Phytoplasma asteris” and “Candidatus Phytoplasma mali” strains infecting sweet and sour cherry in the Czech Republic. Journal of Phytopathology 166: 59–66. doi: 10.1111/jph.12661
Frisinghelli C, Delaiti L, Grando MS, et al. (2000) Cacopsylla costalis (Flor 1861), as a vector of apple proliferation in Trentino. J. Phytopathology 148: 425-431. doi: 10.1046/j.1439-0434.2000.00403.x
Galetto L, Bosco D, Marzachi C (2005) Universal and group-specific real-time PCR diagnosis of flavescence doree (16Sr-V), bois noir (16Sr-Xii) and apple proliferation (16Sr-X) phytoplasmas from fieldcollected plant hosts and insect vectors. Annals of Applied Biology 147: 191-205. doi: 10.1111/j.1744-7348.2005.00030.x
Grando MS, Forti D, Vindimian ME (1998) DNA sequences of the apple proliferation phytoplasma found in psyllid collected from diseased apple trees. Journal of Plant Pathology 80(3): 257.
Hashemi‐Tameh M, Bahar M, Zirak L (2014) Molecular Characterization of Phytoplasmas Related to Apple Proliferation and Aster Yellows Groups Associated with Pear Decline Disease in Iran. Journal of Phytopathology 162: 660-669. doi: 10.1111/jph.12245
Hogenhout SA, Oshima K, Ammar el-D, et al. (2008) Phytoplasmas: bacteria that manipulate plants and insects. Molecular Plant Pathology 9(4): 403–423. doi: 10.1111/j.1364-3703.2008.00472.x
Huang W, MacLean AM, Sugio A, et al. (2021) Parasitic modulation of host development by ubiquitin-independent protein degradation. Cell 184: 1–14. doi: 10.1016/j.cell.2021.08.029
Janik K, Mithöfer A, Raffeiner M, et al. (2017) An effector of apple proliferation phytoplasma targets TCP transcription factors—a generalized virulence strategy of phytoplasma?. Molecular Plant Pathology 18: 435-442. doi: 10.1111/mpp.12409
Jarausch W, Saillard C, Dosba F, Bove J-M (1994) Differentiation of mycoplasmalike organisms (MLOs) in European fruit trees by PCR using specific primers derived from the sequence of a chromosomal fragment of the apple proliferation MLO. Applied and Environmental Microbiology 60(8): 2916-2923. Link
Jarausch W, Saillard C, Helliot B, et al. (2000) Genetic variability of apple proliferation phytoplasmas as determined by PCR-RFLP and sequencing of a non-ribosomal fragment. Molecular and Cellular Probes 14: 17-24. doi: 10.1006/mcpr.1999.0279
Jarausch B, Schwind N, Jarausch W, Krczal G (2003) First report of Cacopsylla picta as a vector of apple proliferation phytoplasma in Germany. Plant Disease 87(10): 101. doi: 10.1094/PDIS.2003.87.1.101A
Jarausch W, Peccerella T, Schwind N, et al. (2004) Establishment of a quantitative real-time PCR assay for quantification of apple proliferation phytoplasmas in plants and insects. Acta Horticulturae 657: 415-420. doi: 10.17660/ActaHortic.2004.657.66
Jarausch B, Schwind N, Fuchs A, Jarausch W (2011) Characteristics of the spread of apple proliferation by its vector Cacopsylla picta. Phytopathology 101: 1471-1480. doi: 10.1094/PHYTO-01-11-0012
Kaminska M, Sliwa H (2003) Effect of antibiotics on the symptoms of stunting disease of Magnolia liliflora plants. Journal of Phytopathology 151: 59-63. doi: 10.1046/j.1439-0434.2003.00682.x
Kaminska M, Sliwa H (2004) First report of phytoplasma belonging to apple proliferation group in roses in Poland. Plant Disease 88: 1283. doi: 10.1094/PDIS.2004.88.11.1283A
Kaminska M, Sliwa H (2008) First report of ‘Candidatus Phytoplasma mali’ in oriental lilies and its association with leaf scorch in Poland. Plant Pathology 57: 363. doi: 10.1111/j.1365-3059.2007.01731.x
Kitazawa Y, Iwabuchi N, Maejima K, et al. (2022) A phytoplasma effector acts as a ubiquitin-like mediator between floral MADS-box proteins and proteasome shuttle proteins, The Plant Cell koac062. doi: 10.1093/plcell/koac062
Krczal G, Bliefernich K (1992) Distribution of apple proliferation disease in orchards with integrated and conventional pesticide programs. Acta Horticultura, 309:253-260.
Krczal G, Krczal H, Kunze L (1988) Fiebierella florii (Stsl), a vector of apple proliferation agent. Acta Horticulturae 235: 99-106.
Krizanac I, Mikec I, Budinscak Z, et al. (2010) Diversity of phytoplasmas infecting fruit trees and their vectors in Croatia. Journal of Plant Diseases and Protection 117(5): 206-213. doi: 10.1007/BF03356362
Kube M, Schneider B, Kuhl H, et al. (2008) The linear chromosome of the plant-pathogenic mycoplasma ‘Candidatus Phytoplasma mali’. BMC Genomics 9: 306. doi: 10.1186/1471-2164-9-306
Kunze L (1979) Damage of root system caused by apple proliferation. Mitteilungen aus der Biologischen Bundesanstalt für Land-und Forstwirtschaft Berlin-Dahlem, 191:204-205.
Lauer U, Seemüller E (2000) Physical map of the chromosome of the apple proliferation phytoplasma. Journal of Bacteriology 182(5): 1415-1418. doi: 10.1128/JB.182.5.1415-1418.2000
Lee IM, Hammond RW, Davis RE, Gundersen DE (1993) Universal amplification and analysis of pathogen 16S rDNA for classification and identification of mycoplasmalike organisms. Phytopathology 83(8): 834-842. doi: 10.1094/Phyto-83-834
Lee IM, Bertaccini A, Vibio M, Gunderson DE (1995) Detection of multiple phytoplasmas in perennial fruit trees with decline symptoms in Italy. Phytopathology 85: 728-735. doi: 10.1094/Phyto-85-728
Lemmetty A, Soukainen M, Tuovinen T (2013) First report of ‘Candidatus Phytoplasma mali’, the causal agent of apple proliferation disease, in apple trees in Finland. Plant Disease doi: 10.1094/PDIS-04-13-0397-PDN
Loi N, Ermacora P, Carraro L, et al. (1988) Apple proliferation detection using monoclonal antibodies. In: XII International Organisation of Mycoplasmology Abstracts, 22-28 July. Australian Microbiological Society/International Organisation for Mycoplasmology: Sydney, Australia, 73-74.
Loi N, Ermacora P, Carraro L, et al. (2002) Production of monoclonal antibodies against apple proliferation phytoplasma and their use in serological detection. European Journal of Plant Pathology 108: 81-86. doi: 10.1023/A:1013901706383
Marcone C, Neimark H, Ragozzino A, et al. (1999) Chromosome sizes of phytoplasmas composing major phylogenetic groups and subgroups. Phytopathology 89: 805‐810. doi: 10.1094/PHYTO.1999.89.9.805
Mehle N, Brzin J, Boben J, et al. (2007) First report of ‘Candidatus Phytoplasma mali’ in Prunus avium, P. armeniaca and P. domestica. Plant Pathology 56: 721-721. doi: 10.1111/j.1365-3059.2007.01597.x
Monti M, Martini M, Tedeschi R (2013) EvaGreen Real-time PCR protocol for specific ‘Candidatus Phytoplasma mali’ detection and quantification in insects. Molecular and Cellular Probes 27(3-4):129-36. doi: 10.1016/j.mcp.2013.02.001
Musetti R, Sanita di Toppi L, Ermacora P, Favali M (2004) Recovery in apple trees infected with the apple proliferation phytoplasma: an ultrastructural and biochemical study. Phytopathology 94: 203-208. doi: 10.1094/PHYTO.2004.94.2.203
Musetti R, Paolacci A, Ciaffi M, et al. (2010) Phloem cytochemical modification and gene expression following the recovery of apple plants from apple proliferation disease. Phytopathology 100: 390-399. doi: 10.1094/PHYTO-100-4-0390
Németh M (1986) Virus, Mycoplasma and Rickettsia Diseases of Fruit Trees. Lancaster, Boston, USA/Dordrecht, Netherlands: M. Nijhoff Publishers, 841 pp.
Osler R, Petrovic N, Ermacora P, et al. (2001) Control strategies of apple proliferation, a serious disease occurring both in Slovenia and Italy. 5th Slovenian Conference on Plant Protection, Catez ob Savi (Slovenia), 6-8 March 2001. Pp. 238-243.
Paltrinieri S, Duduk B, Dal Molin F, et al. (2010) Molecular characterization of ‘Candidatus Phytoplasma mali’ strains in outbreaks of apple proliferation in north eastern Italy, Hungary, and Serbia. Julius-Kühn-Archiv 427: 178-182.
Pedrazzoli F, Filippi M, Deromedi M, et al. (2008) Apple proliferation transmission by grafting in different periods of the year. Acta Horticulturae 781: 489-493. doi: 10.17660/ActaHortic.2008.781.71
Pedrazzoli F, Ciccotti AM, Bianchedi PL, et al. (2008) Seasonal colonization behavior of ‘Candidatus Phytoplasma mali’ in apple trees in Trentino. Acta Horticulturae 781: 483-488. doi: 10.17660/ActaHortic.2008.781.70
Perilla-Henao LM, Casteel CL (2016) Vector-Borne Bacterial Plant Pathogens: Interactions with Hemipteran Insects and Plants. Frontiers in Plant Science 7: 1163. doi: 10.3389/fpls.2016.01163
Rekab D, Pirajno G, Cettul E, et al. (2010) On the apple proliferation symptom display and the canopy colonization pattern of ‘Candidatus Phytoplasma mali’ in apple trees. European Journal of Plant Pathology 127: 7-12. doi: 10.1007/s10658-010-9579-z
Rumbou A, Carraro L, Nanos G, et al. (2008) First report of ‘Candidatus Phytoplasma mali’ in Greece and correlation with small apple fruit disorder occurring in the orchards of the Pelion Mountain. Acta Horticulturae 781: 505-509. doi: 10.17660/ActaHortic.2008.781.74
Schaper U, Seemüller E (1982) Condition of the phloem and the persistence of mycoplasmalike organisms associated with apple proliferation and pear decline. Phytopathology 72: 736-742.
Seemüller E (1976) Investigation to demonstrate mycoplasmalike organisms in diseased plants by fluorescence microscopy. Acta Hort 67: 109-112.
Seemüller E (1990) Apple proliferation. In: Compendium of apple and pear diseases. St Paul, Minnesota, USA: American Phytopathological Society, 67-68.
Seemüller E, Schneider B, Maurer R, et al. (1994) Phylogenetic classification of phytopathogenic mollicutes by sequence analysis of 16S ribosomal DNA. International Journal of Systematic Bacteriology 44(3): 440-446.
Seemüller E, Marcone C, Lauer U, Ragozzino A, Goschl M (1998) Current status of molecular classification of the phytoplasmas. Journal of Plant Pathology 80(1): 3-26. Link
Seemüller E, Schneider B (2004) ‘Candidatus Phytoplasma mali’, ‘Candidatus Phytoplasma pyri’, and ‘Candidatus Phytoplasma prunorum’, the causal agents of apple proliferation, pear decline, and European stone fruit yellows, respectively. International Journal of Systemic and Evolutionary Microbiology 54: 1217-1226. doi: 10.1099/ijs.0.02823-0
Seemüller E, Schneider B (2007) Differences in virulence and genomic features of strains of ‘Candidatus Phytoplasma mali’ the apple proliferation agent. Phytopathology 97: 964-970. doi: 10.1094/PHYTO-97-8-0964
Seemüller E, Kiss E, Sule S, Schneider B (2010) Multiple infection of apple trees by distinct strains of ‘Candidatus Phytoplasma mali’ and its pathological relevance. Phytopathology 100: 863-870. doi: 10.1094/PHYTO-100-9-0863
Smart CD, Schneider B, Blomquist CL, et al. (1996) Phytoplasma-specific PCR primers based on sequences of the 16S-23S rRNA spacer region. Applied and Environmental Microbiology 62(8): 2988-2993. Link
, , , et al. (2023) Target degradation specificity of phytoplasma effector phyllogen is regulated by the recruitment of host proteasome shuttle protein. Molecular Plant Pathology 00: 1–12. doi: 10.1111/mpp.13410
Vindimian ME, Ciccotti A, Filippi M, et al. (2002) Spread of apple proliferation by root bridges (Abstr.). Petria 12: 375.
Tedeschi R, Visentin C, Alma A, Bosco D (2003) Epidemiology of apple proliferation (AP) in northwestern Italy: evaluation of the frequency of AP‐positive psyllids in naturally infected populations of Cacopsylla melanoneura (Homoptera: Psyllidae). Annals of Applied Biology 142: 285-290. doi: 10.1111/j.1744-7348.2003.tb00252.x
Tedeschi R, Alama A (2004) Transmission of apple proliferation phytoplasma by Cacopsylla melanoneura (Homoptera: Psyllidae). Journal of Economic Entomology 97(1):8-13. Link
Tedeschi R, Alma A (2006) Fieberiella florii (Homoptera: Auchenorrhyncha) as a vector of ‘Candidatus Phytoplasma mali’. Plant Disease 90: 284-290. doi: 10.1094/PD-90-0284
Tedeschi R, Lauterer P, Brusetti L, et al. (2009) Composition, abundance and phytoplasma infection in the hawthorn psyllid fauna of northwestern Italy. European Journal of Plant Pathology 123(3): 301–310. doi: 10.1007/s10658-008-9367-1
Torres E, Bertolini E, Cambra M, Montón C, Martín MP (2005) Real-time PCR for simultaneous and quantitative detection of quarantine phytoplasmas from apple proliferation (16SrX) group. Molecular and Cellular Probes 19(5): 334-340. doi: 10.1016/j.mcp.2005.06.002
Valasevich N, Schneider B (2016) Detection, Identification, and Characterization of Phytoplasmas Infecting Apple and Pear Trees in Belarus. Plant Disease 100(11): 2275-2280. doi: 10.1094/PDIS-04-16-0498-RE
Vega FE, Davis RE, Barbosa P, Dally EL, Purcell AH, Lee IM (1993) Detection of a plant pathogen in a nonvector insect species by the polymerase chain reaction. Phytopathology 83(6): 621-624. doi: 10.1094/Phyto-83-621
, , , et al. (2024) Phytoplasma: A plant pathogen that cannot be ignored in agricultural production—Research progress and outlook. Molecular Plant Pathology 25: e13437. doi: 10.1111/mpp.13437
Weintraub PG, Beandland L (2006) Insect vectors of phytoplasmas. Annual Review of Entomology 51: 91-111. doi: 10.1146/annurev.ento.51.110104.151039
Zimmermann MR, Schneider B, Mithöfer A, Reichelt M, Seemüller E, Furch AC (2015) Implications of Candidatus Phytoplasma mali infection on phloem function of apple trees. Endocytobiosis and cell research: journal of the International Society of Endocytobiology 26: 67-75. Link