Sautua Francisco & Carmona Marcelo
.
El sistema de defensa basal o constitutivo, anteriormente descripto, es inespecífico. Sin embargo, cada célula vegetal posee un sofisticado “sistema de vigilancia” que puede censar y distinguir una gran cantidad de señales de diverso origen, producidas por microorganismos y por el ambiente, permitiéndoles reconocer agentes extraños y activar sus mecanismos de defensa. De esta manera, se inducen en forma rápida y eficiente las defensas en el sitio de infección, creando condiciones adversas para el crecimiento y reproducción de patógenos (Heath, 2000). Los elicitores específicos e inespecíficos parecen desencadenar cascadas de transducción de señales que involucran proteínas quinasas y proteínas fosfatasas. Una proteína quinasa es una enzima que opera como transductor de señales y esto lo hace modificando otras proteínas (sustratos), mediante la fosforilación. Este sistema de señalización hace posible que las plantas expresen toda una batería de diferentes mecanismos de defensa, compuestos por: reforzamiento de las barreras físicas, reacciones químicas (producción de substancias tóxicas) y bioquímicas (enzimas líticas), que inhiben la penetración o el establecimiento de la relación de parasitismo, etc. (Dangl y Jones, 2001).
Entre los mecanismos de respuesta local se encuentran por ejemplo el enriquecimiento de la pared celular con lignina (lignificación), suberina, calosa y glicoproteínas ricas en hidroxiprolina (GRHP), la formación de papilas, o la oclusión vascular. Junto al desarrollo de estructuras locales para bloquear la entrada de los patógenos, alrededor del lugar de la infección también se acumulan sustancias conocidas como fitoalexinas, que incluyen una gran cantidad de productos naturales que poseen capacidad antimicrobiana, y que se sintetizan en mayor cantidad tras la entrada de un patógeno (Hammerschmidt, 1999).
Después de la infección por un patógeno, la colonización de las raíces con ciertos microorganismos beneficiosos o después del tratamiento con diversos productos químicos, muchas plantas establecen una situación fisiológica única que se denomina estado «activado» (del inglés “primed”) de la planta. En este estado, las plantas pueden «recordar» la infección previa, la colonización de la raíz o el tratamiento químico. Como consecuencia, las plantas o tejidos sensibilizados responden de manera más rápida y/o efectiva cuando son reexpuestas al estrés biótico o abiótico, una característica que frecuentemente se asocia con una mayor resistencia a las enfermedades (Goellner y Conrath, 2008).
Muchas plantas y semillas contienen proteínas que inhiben específicamente las enzimas patógenas y plagas al formar complejos que bloquean los sitios activos o alteran las conformaciones enzimáticas, reduciendo finalmente la función enzimática. Estas proteínas son generalmente pequeñas y ricas en el aminoácido cisteína. Incluyen defensinas, inhibidores de amilasa, lectinas e inhibidores de proteinasas. A diferencia de los productos químicos simples como los terpenoides, los compuestos fenólicos y los alcaloides, las proteínas requieren una gran cantidad de recursos y energía para ser producidas; en consecuencia, muchas proteínas defensivas solo se producen en cantidades significativas después de que un patógeno o plaga ha atacado la planta. Sin embargo, una vez activadas, las proteínas y enzimas de defensa inhiben eficazmente hongos, bacterias, nematodos y herbívoros insectívoros.
.
Percepción y transducción de señales de plantas ante patógenos. Elicitores y su mecanismo de acción
La interacción de los sistemas de señalización en las interacciones planta-patógeno es un componente clave en el desarrollo de enfermedades en las plantas. La señalización entre hongos patógenos y plantas comienza cuando se encuentran en la superficie de la planta (Vidhyasekaran, 2007; He et al., 2018). Existe una fuerte evidencia de que las interacciones planta-hongo fitopatógeno comienzan y quizás sus resultados (susceptibilidad o resistencia) se determinan durante los primeros minutos de asociación y tal vez incluso durante un tiempo aún más breve (Heath, 2000). Si las señales en la superficie de la planta son percibidas como favorables por los hongos (reconocimiento de sus hospedantes), sus esporas germinan y penetran directamente en la cutícula o penetran la cutícula produciendo apresorio y cojinete de infección (Nielsen et al, 2000; Wright et al, 2000; Apoga et al. al, 2001; Lev y Horwitz, 2003; Fujita et al, 2004; Li et al, 2004). Las señales físicas y químicas de la planta pueden estar involucradas en el proceso de penetración. Las señales de la planta pueden ser utilizadas por el hongo para desencadenar la expresión de los genes necesarios para penetrar en el hospedante (Dickman et al, 2003, Tsuji et al, 2003, Yang et al, 2005).
A su vez, las plantas han desarrollado sistemas sofisticados de detección y respuesta que descifran las señales de los patógenos e inducen la síntesis de diversos compuestos antifúngicos y fortifican sus componentes de la pared celular para defenderse. Para lograr una infección exitosa, los patógenos deben superar estos mecanismos de defensa produciendo señales apropiadas. En la percepción de señales de patógenos, se activan varias rutas de señalización en plantas que aumentan la respuesta de resistencia a patógenos invasores (Glazebrook et al., 2003; Nandi et al., 2003; Hammerschmidt, 2004; Thaler et al 2004; Uppalapati et al, 2004). La interacción de las señales de plantas y patógenos permite a la planta ajustar las respuestas de defensa (Mould et al, 2003) y el patógeno para sucumbir al ambiente tóxico en las plantas o superar estos mecanismos de defensa, lo que da lugar a susceptibilidad o resistencia a enfermedades (Yang et al, 2004). La susceptibilidad y la resistencia son lados opuestos de la misma moneda e indican el candidato ganador en la batalla entre la planta y el patógeno (Neu et al, 2003; Christensen et al, 2004). La resistencia y la susceptibilidad a menudo se determinan a nivel de célula individual (Gjetting et al, 2004).
.
Qué son los elicitores?
El término “elicitor” fue usado por primera vez para describir moléculas que eran capaces de provocar la producción de fitoalexinas (Keen et al., 1972; Mayer & Ziegler, 1988; Yamaoka et al., 1990; Vidhyasekaran, 1988), pero ahora se usa comúnmente para compuestos que provocan cualquier tipo de defensa de plantas (Boller & Felix, 2009). El término elicitor incluye tanto sustancias de origen patógeno (elicitores exógenos, elicitores microbianos) como compuestos liberados de plantas (elicitores endógenos, elicitores de plantas hospedantes) (Boller, 1995; D’Ovidio et al., 2004).
Los elicitores microbianos se pueden clasificar en dos grupos: elicitores generales y elicitores específicos de raza. Los elicitores generales son los elicitores microbianos que pueden desencadenar respuestas de defensa tanto en plantas hospedantes como no hospedantes (Kruger et al., 2003; Vidhyasekaran, 2007; Lee et al., 2017; Panstruga et al., 2020). Los elicitores específicos de raza inducen respuestas de defensa que conducen a la resistencia a enfermedades solo en cultivares hospedantes específicos (Bonas y Lhayae, 2002; Montesano et al., 2003). Un par complementario de genes en una raza de patógeno particular y un cultivar hospedante determina esta resistencia específica del cultivar (gen por gen). Por lo tanto, un inductor específico de raza codificado o producido por la acción de un gen de avirulencia (avr) presente en una raza particular de un patógeno provoca resistencia solo en una variedad de planta hospedante que porta el gen de resistencia (R) correspondiente (Lu y Higgins, 1998; Cohn et al., 2001; Luderer y Joosten, 2001; Nimchuk et al., 2001; Nurnberger y Scheel, 2001).
Los elicitores fúngicos no tienen ninguna estructura química común y pertenecen a diferentes tipos de clases químicas, tales como oligosacáridos, proteínas, péptidos, glicoproteínas y lípidos. Los elicitores son producidos por patógenos necrotrofos (Desjardins y Plattner, 2000), hemibiotróficos (Yamaoka et al. , 1990) y biotróficos (parásitos obligados) (Ryerson y Heath, 1992). También se han detectado moléculas inductoras en algunos saprófitos (Dean y Anderson, 1991) y en levaduras (Klusener y Weiler, 1999).
Las plantas pueden detectar moléculas microbianas a través de receptores de reconocimiento que se encuentran en la superficie de la célula vegetal y reconocen patrones moleculares microbianos generales (PRR, pattern-recognition receptors); y receptores intracelulares de la familia de receptores que poseen repeticiones ricas en leucina (NLR, leucine-rich repeat receptor) que se unen a nucleótidos que reconocen proteínas efectoras específicas de los patógenos. Las correspondientes respuestas de inmunidad desencadenadas por patrones moleculares (PTI, pattern-triggered immunity) y por efectores (ETI, effector-triggered immunity) se consideraron durante mucho tiempo caminos separados, aunque actualmente se está empezando a comprender que ambas vías convergen en respuestas celulares muy similares, como el flujo de iones de calcio y la superposición de genes de reprogramación. Una serie de estudios descubrieron recientemente la genética e interconexiones moleculares entre PTI y ETI, destacando la complejidad de la red inmune de las plantas. Notablemente, las respuestas inmunitarias mediadas por PRR y NLR requieren y se potencian entre sí para alcanzar un resultado inmunitario óptimo (Bernoux et al., 2022).
.
El modelo actual del sistema inmune de las plantas, de reconocimiento de patógenos y activación de defensas
Recientemente, la convergencia entre estudios moleculares sobre la inmunidad vegetal y las estrategias de infección de patógenos están comenzando a revelar una visión integrada de la interacción planta-patógeno desde la perspectiva de ambos organismos. Las plantas tienen una capacidad asombrosa para reconocer patógenos a través de estrategias que implican elicitores (inductores) de patógenos tanto conservados o inespecíficos como variables u específicos, y a su vez los patógenos manipulan la respuesta de defensa a través de la secreción de moléculas efectoras de virulencia.
Las plantas han desarrollado múltiples capas de sofisticados mecanismos de vigilancia que reconocen los patógenos potencialmente peligrosos y responden rápidamente antes de que esos organismos tengan la oportunidad de causar un daño grave. Estos sistemas de vigilancia están vinculados a respuestas de defensa preprogramadas específicas.
El modelo actual en el cual se basa nuestra comprensión del funcionamiento del sistema inmune de las plantas es el modelo de “zigzag” propuesto por Jones y Dangl (2006). Este modelo ilustra la expresión cuantitativa del sistema inmunológico de las plantas, en cuatro fases diferentes (Figura 1).
.
-
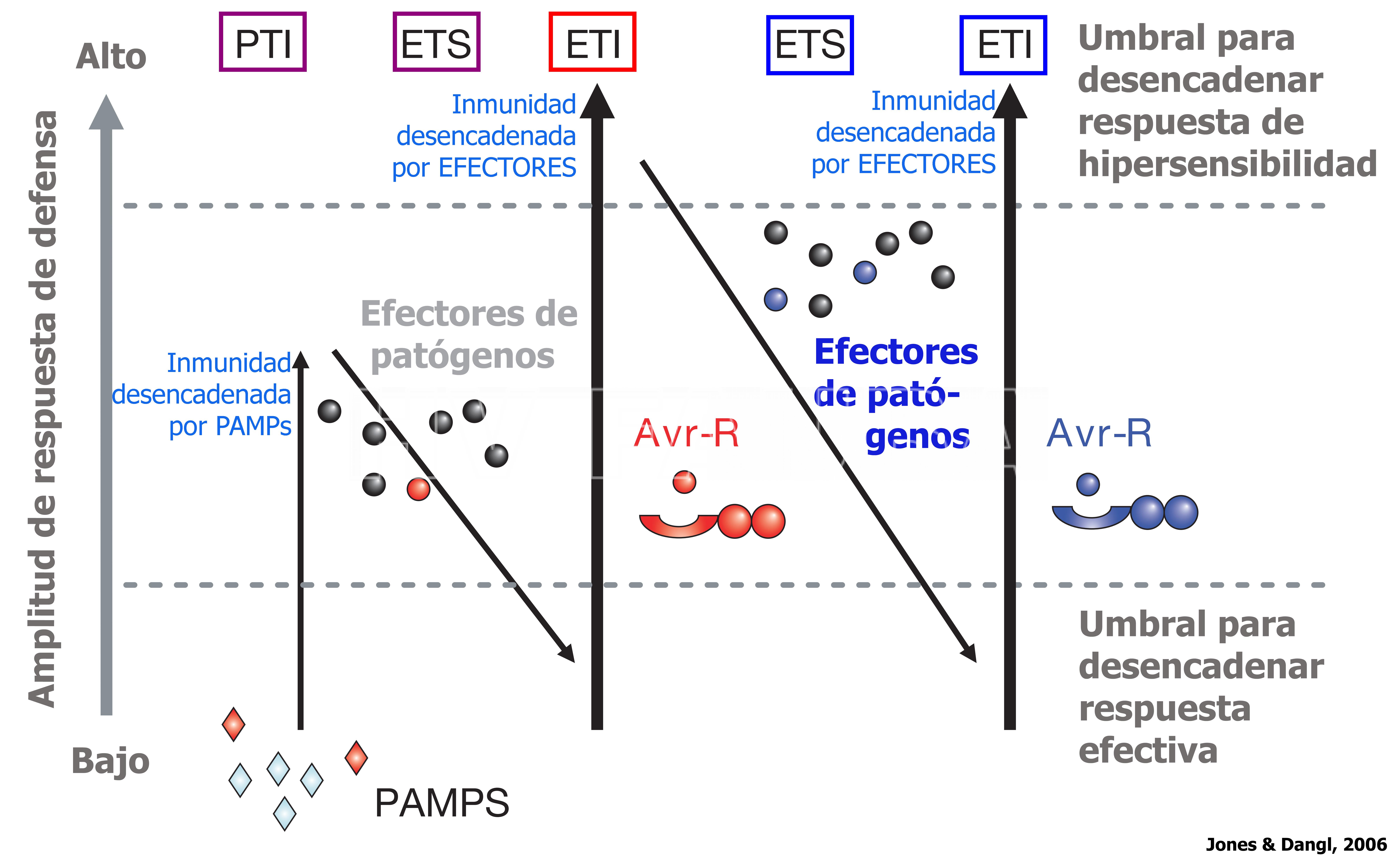
- Fig. 1. Representación conceptual del Modelo de inmunidad de las plantas denominado de “zigzag”, propuesto por Jones y Dangl (2006).
Figura 1. Modelo de “zigzag” propuesto por Jones y Dangl (2006). En la fase 1, los PAMPs son reconocidos por los PRRs, lo que resulta en la activación de la PTI, que puede detener la colonización adicional del tejido vegetal. En la fase 2, los patógenos exitosos liberan efectores que contribuyen a la virulencia, los que pueden interferir con la PTI, y esto resulta en una ETS (del inglés: effector-triggered susceptibility). En la fase 3, determinados efectores son reconocidos por una de las proteínas de resistencia (R) (ej. NB-LRR), resultando en la activación de la ETI, la cual es una respuesta acelerada y amplificada de PTI, lo que conlleva a la resistencia a la enfermedad y casi siempre, a una respuesta de muerte celular programada en el sitio de la infección. En la fase 4, la selección natural conduce al patógeno a evitar la ETI eliminando o diversificando el gen efector reconocido o adquiriendo determinantes antigénicos o efectores adicionales, que les permiten evadir la ETI. A su vez, en las plantas la selección natural da lugar a nuevas especificidades R, que deriva en el accionar la ETI nuevamente.
.
El primer paso en la defensa de las plantas consiste en detectar la presencia de patógenos que intentan penetrar sus tejidos e infectar. Para reconocer y prevenir la infección por virus, bacterias, oomycetes, hongos, nemátodos, etc., las plantas han desarrollado evolutivamente dos estrategias que consisten en esencia en dos sistemas principales de receptores inmunes (Jones y Dangl, 2006; Pritchard & Birch, 2014).
.
.
INMUNIDAD ACTIVADA POR PAMP (PTI)
El primer sistema involucra receptores ubicados en la superficie celular (transmembrana), denominados “receptores de reconocimiento de patrones moleculares» (PRRs, del inglés: pattern recognition receptors) (Tör et al., 2009; Kagan, 2023). Es decir, sobre la faz externa de las células vegetales, elicitores microbianos conservados, denominados “patrones moleculares asociados a patógenos” (PAMPs, del ingles: pathogen-associated molecular patterns), son reconocidos por proteínas receptoras denominadas PRPs. Inicialmente, las plantas detectan los PAMPs a través de los mencionados receptores PRRs, ubicados en la superficie celular (fase 1 del modelo de zigzag). Los PAMPs son moléculas que evolutivamente se han conservado en las diferentes especies y son características de grupos enteros de microorganismos, como por ejemplo las flagelinas en las bacterias, quitina en los hongos, lipopolisacáridos, etc. (Zipfel 2008; Zipfel, 2014; Zhong et al., 2017; Khokhani et al., 2021). Los PAMPs reconocidos por las plantas son diversísimos e incluyen proteínas, carbohidratos, lípidos y moléculas pequeñas como por ejemplo el ATP (Dodds & Rathjen, 2010). De esta manera, las plantas han evolucionado para reconocer características comunes de los patógenos microbianos. Por esta razón, a esta resistencia también se la denomina desencadenada por “patrones moleculares asociados a microorganismos” (MAMPs, del inglés: microbial-associated molecular patterns). Las plantas también responden a moléculas endógenas liberadas a partir de la invasión de patógenos, tales como fragmentos de pared celular o cutícula, denominados “patrones moleculares asociados a peligro” (DAMPs, del inglés: danger-associated molecular patterns). De esta manera, PAMPs, MAMPs y DAMPs son percibidos por receptores de reconocimiento transmembrana, que activan la resistencia o los mecanismos de defensa, después de la detección (Boller & Felix, 2009). Los PAMPs son definidos como epítopos invariantes dentro de moléculas conservadas, indispensables para los microorganismos patógenos, ausentes en plantas y reconocidos por un gran número de hospedantes potenciales, por lo tanto es difícil su transformación o supresión. Esta percepción desencadena la denominada “inmunidad desencadenada por patrones moleculares asociados a patógenos” (PTI, del inglés: PAMP-triggered immunity).
Las plantas detectan inicialmente los microorganismos a través de la percepción de los PAMPs mediante sus PRRs situados en la superficie celular (Zipfel, 2008; Monaghan & Zipfel, 2012; Zipfel, 2014;). Este primer nivel de reconocimiento, como se refirió anteriormente, es el asociado con la PTI, cuya importancia se hizo evidente por la gran susceptibilidad a bacterias que presentan mutantes de PRR o de los componentes de la PTI. Uno de los ejemplos más estudiados es el reconocimiento de la proteína flagelina, la cual forma los filamentos de las bacterias flageladas, por el receptor quinasa “sensor de flagelina 2” (FLS2, del inglés: flagelin sensing 2). La flagelina se liga directamente al receptor FLS2 desencadenando una activa y compleja señalización. Otro ejemplo de PTI lo constituye la regulación estomática. Recientemente se han identificado MAMPs en omycetes (Kato et al., 2021; Monjil et al., 2021).
.
PRR
Los PRRs tienen gran afinidad y especificidad, son capaces de reconocer un PAMP específico a concentraciones por debajo de los nanomoles (Zipfel 2008; Zipfel, 2014). Receptores de MAMP para ciertas proteínas y fragmentos de quitina de hongos han sido identificados como receptores de superficie sin dominio quinasas en plantas (Tör et al., 2009; He et al., 2018).
.
Señalización del reconocimiento PAMP-PRR
Los PRRs, por sí mismos, no pueden emitir la señal al interior de la célula. El reconocimiento de los PAMPs acciona diversos cambios moleculares y fisiológicos. Dentro de segundos y minutos, después de este reconocimiento, se incrementa el flujo de iones a través de la membrana plasmática, y aumenta notablemente la concentración de Ca2+ (Gao et al., 2021), se activan las proteínas MAPK, ocurre la fosforilación de varias proteínas, la producción de especies reactivas del oxígeno, la endocitosis del receptor e interacciones proteínas-proteínas, y a los 30 minutos se inducen los cambios transcripcionales de determinados genes.
Estos genes relacionados con la defensa se activan mediante dos vías de señalización, una mediada por el MeJA/Et (del inglés: methyl jasmonate/ethylene, hormonas metil jasmonato y etileno), y otra por el ácido salicílico (SA), otra hormona vegetal, lo que indica que el mismo PAMP puede activar múltiples cascadas de señalización (Fernández-Milmanda, 2023).
Las respuestas intracelulares asociadas a la PTI incluyen una rápida difusión de iones a través de la membrana plasmática, la activación de la cascada de proteínas MAPK (del inglés: mitogen-activated protein kinase), la producción de especies reactivas del oxígeno, rápidos cambios en la expresión de genes y un reforzamiento de la pared celular.
.
.
Supresión de la PTI por efectores patogénicos
A pesar de la efectividad de todos estos mecanismos moleculares que se han descrito anteriormente, existen muchos patógenos exitosos que han desarrollado factores de virulencia para evadir o suprimir la PTI, como pueden ser, incluyendo fitotoxinas, polisacáridos extracelulares y efectores proteicos.
En el caso de las bacterias, la mayoría de estas moléculas se secretan mediante el sistema de secreción tipo III, denominado TTSS o T3SS (del inglés: type III secretion system). Las bacterias patógenas de plantas pueden introducir de 15 a 30 efectores dentro de las células del hospedante usando el TTSS. Los efectores bacterianos contribuyen a la virulencia del patógeno, ya sea mimetizando o inhibiendo las funciones celulares. No todas las bacterias, incluso no todas las cepas de P. syringae, expresan estas proteínas, lo cual sugiere que existen otras estrategias para inhibir las señales de los PRRs. Otros tipos de efectores atacan directamente los procesos que están por debajo de la cascada de señalización, y traen otras consecuencias.
Efectores de hongos y de oomycetes pueden actuar tanto en la matriz extracelular o en el interior de la célula del hospedante (Fabro, 2021; Pi et al., 2021). Otros, a su vez, actúan probablemente dentro de la célula del hospedante y son reconocidos por las proteínas de NB-LRR. Aun se desconoce cómo estos efectores de hongos y oomycetes son liberados al interior de la célula y cómo contribuyen a la virulencia.
En el caso de los nemátodos, el efector del nematodo del quiste parásito GLAND4 es una proteína de unión al ADN. Los nematodos del quiste de la remolacha azucarera y la soja poseen el efector GLAND4, el cual tiene capacidad de unión al ADN y reprime la expresión de un gen “informador”. Este efector nuclear es el primer efector reportado de nematodos parásiticos de unión al ADN (Barnes et al., 2018).
.
INMUNIDAD ACTIVADA POR EFECTORES PATOGÉNICOS (ETI)
Durante el proceso evolutivo, una vez que los patógenos adquirieron la capacidad de suprimir las defensas primarias, las plantas desarrollaron un segundo mecanismo de reconocimiento especializado para detectarlos. Este sistema de reconocimiento de patógenos es controlado por los genes de resistencia mayores del hospedante. En este sistema inmune denominado «gen por gen», los genes de resistencia (R) de la planta confieren resistencia contra el patógeno, quien a su vez contiene los correspondientes genes de avirulencia (Avr) respectivos. Los eventos de reconocimiento implican que los productos de los genes R y Avr interactúen desencadenando las respuestas de la defensa del hospedante, incluyendo la muerte localizada de la célula hospedante o una respuesta hipersensible (HR) que limita la propagación del patógeno (Balint-Kurti, 2019). Se denominan genes de avirulencia porque cuando están presentes en el genoma del patógeno, el hospedante los puede detectar (siempre que a su vez posea el gen de resistencia correspondiente) previniendo el desarrollo de la enfermedad. Cuando las plantas reconocen la presencia de las moléculas efectoras de patógenos por medio de las proteínas de resistencia, se desencadena la inmunidad activada por efectores (ETI, del inglés: effector-triggered immunity) (Gassmann & Bhattacharjee, 2012).
Los patógenos y simbiontes de las plantas secretan proteínas denominadas efectores para facilitar la infección al suprimir las respuestas de defensa de las plantas y alterar la estructura y función de la célula hospedante (Sperschneider y Dodds, 2022). Los patógenos secretan efectores en el espacio extracelular del hospedante (el apoplasto de la planta) así como en el espacio intracelular (el citoplasma de la planta). Los efectores apoplásicos pueden funcionar en el apoplasto (De Wit, 2016) o pueden unirse a la pared celular del hongo (Tanaka y Kahmann, 2021). Por el contrario, los efectores citoplásmicos se introducen dentro de la célula vegetal y, posteriormente, algunos pueden dirigirse a compartimentos celulares específicos de la planta. Los efectores citoplasmáticos tienen una mayor proporción de aminoácidos cargados positivamente, mientras que los efectores apoplásticos están enriquecidos con residuos de cisteína. En el caso de pseudohongos como por ejemplo Phytophthora infestans, los haustorios son sitios para el suministro y secreción de efectores tanto apoplásticos como citoplasmáticos durante la infección, siguiendo distintas vías de secreción (Wang et al., 2017). La entrega de efectores en las células hospedantes está bien caracterizada en bacterias, nematodos e insectos (Galán et al., 2014; Mitchum et al., 2013; Rodriguez y Bos, 2013). Por ejemplo, las bacterias tienen sistemas de secreción conservados, como el sistema de secreción de tipo III, para inyectar efectores directamente en la célula vegetal, y los nematodos usan un estilete para inyectarlos efectores en la célula hospedante e infectar. Sin embargo, los mecanismos de entrega de efectores en hongos y oomicetes siguen siendo difíciles de caracterizar (Lo Presti y Kahmann, 2017). En Magnaporthe oryzae, los efectores citoplasmáticos se acumulan preferentemente en el complejo interfacial biotrófico (Khang et al., 2010), mientras que los efectores apoplásicos siguen la vía secretora convencional (Giraldo et al., 2013). En Ustilago maydis, se ha sugerido que un complejo proteico estable de cinco efectores y dos proteínas de membrana está involucrado en la translocación del efector; sin embargo, estas proteínas no se conservan ampliamente en los hongos (Ludwig et al., 2021). La presencia de motivos de secuencia conservados en efectores de oomicetes de secuencia divergente se ha implicado en la entrega de efectores (Whisson et al., 2007). El motivo RxLR aparece en la secuencia N-terminal después del sitio de escisión del péptido señal y, a menudo, va seguido de una extensión ácida de aminoácidos. Sin embargo, se ha debatido la validación experimental de la captación del efector mediada por RxLR (Ellis y Dodds, 2011), con evidencia reciente que apunta hacia un papel del motivo RxLR en la escisión del efector antes de la secreción (Wawra et al., 2017). Además, el motivo RxLR no se conserva fuera de Phytophthora spp. y del mildiú. El dominio WY es un dominio estructural asociado con efectores de oomicetes citoplasmáticos del orden Peronosporales que se ha utilizado para identificar candidatos a efectores con motivos RxLR degenerados (Wood et al., 2020). De manera similar, se han aplicado búsquedas de motivos RxLR degenerados en Pythium spp. que carecen de efectores con un motivo RxLR canónico (Ai et al., 2020). El motivo de secuencia LxLFLAK define las proteínas de la familia inductora de arrugas y necrosis (CRN) y también se usa comúnmente para buscar efectores CRN (Schornack et al., 2010).
Los efectores intracelulares, principalmente proteínas pero también pequeños ácidos ribonucleicos, son transportados por los patógenos al citoplasma de la célula hospedante donde interfieren con la fisiología normal de la planta. Los diversos procesos del hospedante afectados incluyen la transcripción de genes y el silenciamiento mediado por ARN, la muerte celular, la estabilidad de proteínas, la secreción de proteínas y la autofagia (Fabro, 2021). Algunos objetivos o target de los efectores están directamente involucrados en la ejecución de las respuestas de defensa, mientras que otros participan en metabolismos fundamentales cuya alteración afecta colateralmente a las defensas. Otros objetivos efectores son los factores de susceptibilidad (SF), es decir, los componentes del hospedante que hacen que las plantas sean vulnerables a los patógenos. Los SF son en su mayoría reguladores negativos de la inmunidad, pero algunos parecen necesarios para mantener o promover la colonización de patógenos.
La mayoría de los genes R conocidos codifican receptores inmunes que son proteínas de tipo NLR o NB-LRR (Monteiro & Nishimura, 2018). Estas proteinas poseen una estructura que contiene un sitio central de unión a nucleótidos (NB, del inglés: nucleotide-binding) de tipo ARC (APAF-1, proteínas de resistencia a enfermedades, CED-4) y un dominio C terminal de repetición rico en leucina (LRR, del inglés: leucine-rich repeat) (Jacob et al. , 2013; Jones et al., 2016). El dominio LRR está implicado en el reconocimiento de los diferentes patógenos. En Arabidopsis thaliana (Arabidopsis), la mayoría de estos resceptores contienen en su extremo N-terminal una hélice superenrollada (CC, del inglés: coiled coil) o un dominio TIR (del inglés: Toll Interleukin-1 Receptor) (Meyers et al., 2003; Eastman et al., 2022; Jia et al., 2023). En general, los genes de CC-NB-LRR están presentes en los genomas de monocotiledóneas y dicotiledóneas, mientras que los genes de TIR-NB-LRR están completamente ausentes en monocotiledóneas, pero están fuertemente representados en dicotiledóneas (Jacob et al., 2013). Ejemplos de otros dominios presentes en la parte N-terminal de los receptores NLR son los dominios de Solanaceae (SD, del inglés: Solanaceae domains) que poseen algunos NLR de plantas de la familia de las Solanaceae, dominios “BED-DNA-binding zinc-finger” de NLRs de Puccinia striiformis f. sp. tritici (Marchal et al., 2018) o de Populus trichocarpa; o los dominios WRKY en las proteínas de Arabidopsis WRKY19 / MEKK4 y RRS1. Aunque los repertorios de NLR son relativamente compactos en linajes de plantas ancestrales, la presencia de estas proteínas se expandió masivamente a una de las familias más grandes y variables en clados de plantas avanzadas, llegando a más de 400 miembros en arroz (Oryza sativa) o uva (Vitis vinifera) (Clark et al., 2007; Jacob et al., 2013; Ossowski et al., 2008). El número de genes NLR parece correlacionarse generalmente con el número total de genes en un genoma. En muchos casos, los NLR pueden reconocer directamente los efectores patógenos uniéndose a ellos. No parece haber reglas generalizables mediante las cuales el dominio NLR se une a los efectores patógenos. Un papel en la unión del sustrato tiene sentido intuitivo dado el papel conocido de los dominios LRR como plataformas de unión a diversos sustratos (Fluhr, 2001). Posteriormente, se ha encontrado que los dominios LRR se unen directamente a diversos efectores y también se encuentran bajo una selección diversificada, presumiblemente impulsada por la diversidad de los efectores y la diversificación (Goritschnig et al., 2016, Krasileva et al., 2010; De la Concepcion et al., 2018). Más allá del dominio LRR, otros dominios también participan claramente en el reconocimiento. En el caso de los genes en el locus L de lino, los NLR de tipo TIR (TNL) con LRR idénticos pero los dominios TIR ligeramente divergentes tienen especificidades distintas (Ellis et al., 1999).
.
-
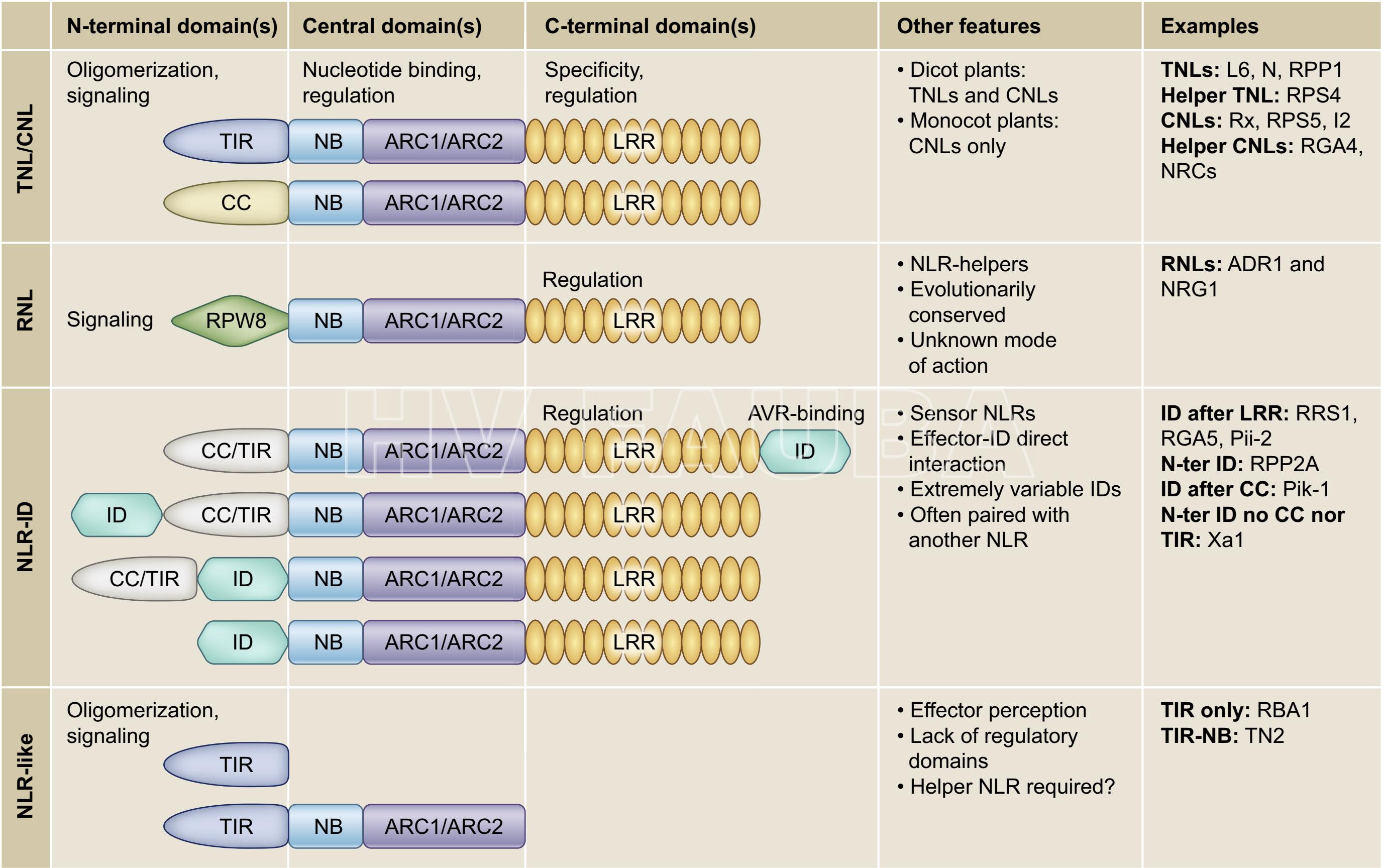
- Fig. 2. Representación esquemática de proteínas de defensa NLR de plantas conocidas (Cesari, 2018).
Figura 2. Representación esquemática de la flexibilidad y diversidad de la arquitectura de proteínas de defensa NLR de plantas conocidas (Cesari, 2018). Se indican las funciones o propiedades predichas y documentadas experimentalmente de dominios individuales, así como características y ejemplos particulares. Los dominios no están dibujados a escala. CC = bobina enrollada o hélice superenrollada (del inglés: coiled-coil); TIR = receptor de peaje / interleuquina-1 (del inglés: toll/interleukin-1 receptor); NB = unión de nucleótidos (del inglés: nucleotide binding); ARC1/2, Apaf-1, productos del gen R y CED4; LRR = repetición rica en leucina (del inglés: leucine-rich repeat); ID = dominio integrado (del inglés: integrated domain); RPW8 = resistencia al oídio 8 (del inglés: resistance to powdery mildew 8); NLR = proteína de unión a nucleótidos y rica en repeticiones de leucina (del inglés: nucleotide-binding and leucine-rich repeat-containing protein); TNL = TIR-NLR; CNL = CC-NLR; RNL = RPW8-NLR.
.
En cambio, las proteínas Avr son diversas, y muchas tienen funciones importantes en el proceso de infección. Los genes avr de patógenos evolucionan rápidamente y codifican proteínas efectoras polimórficas que a menudo son introducidas en las células vegetales por parte de los patógenos, donde interactúan con las proteínas del hospedante para atenuar la respuesta inmune del hospedante o mejorar la aptitud patógena (virulencia) en el ambiente del hospedante (Abramovitch et al., 2006; Chisholm et al., 2006; Dangl y McDowell, 2006). El reconocimiento de los efectores por parte de los receptores inmunes NLR desencadena un conjunto de reacciones de defensa denominadas inmunidad desencadenada por efectores (ETI). Pero, cómo los NLRs inducen la ETI es poco conocido. En este punto, no se identificó una vía general de inducción de defensas mediada por NLR, y parecería que muchos mecanismos diferentes han evolucionado para vincular el reconocimiento de efectores facilitados por NLR con las respuestas de defensa aguas abajo (Dodds and Rathjen, 2010; Li et al., 2015). Además de servir como receptores específicos para los efectores patógenos, se han propuesto que varios miembros ampliamente conservados de esta familia de receptores funcionan como NLR auxiliares, es decir, que tienen una función en la transducción o amplificación de señales de defensa (Bonardi et al., 2011; Li et al. al., 2015). La relación antagónica entre los genes de R y Avr da lugar a conflictos coevolutivos, y la selección favorece la evolución de la resistencia en plantas o la virulencia en los patógenos.
La ETI es, probablemente, una de las formas de mayor alcance en respuestas de defensa en plantas (He et al., 2007; Chiang & Coaker, 2015 ). La mayoría de los genes R codifican para las proteínas NB-LRR (Ayliffe & Lagudah, 2004). Existen aproximadamente 125 en el genoma de A. thaliana. Si un efector es reconocido por la proteína NB-LRR correspondiente, se activa la ETI, que es más rápida y de mayor intensidad de respuesta que la PTI y a menudo culmina en una HR. La HR no se extiende normalmente más allá de la célula infectada: puede retardar el crecimiento del patógeno en algunas interacciones, particularmente las que involucran patógenos formadores de haustorios, pero no siempre se puede observar, ni se requieren, para ETI.
.
-
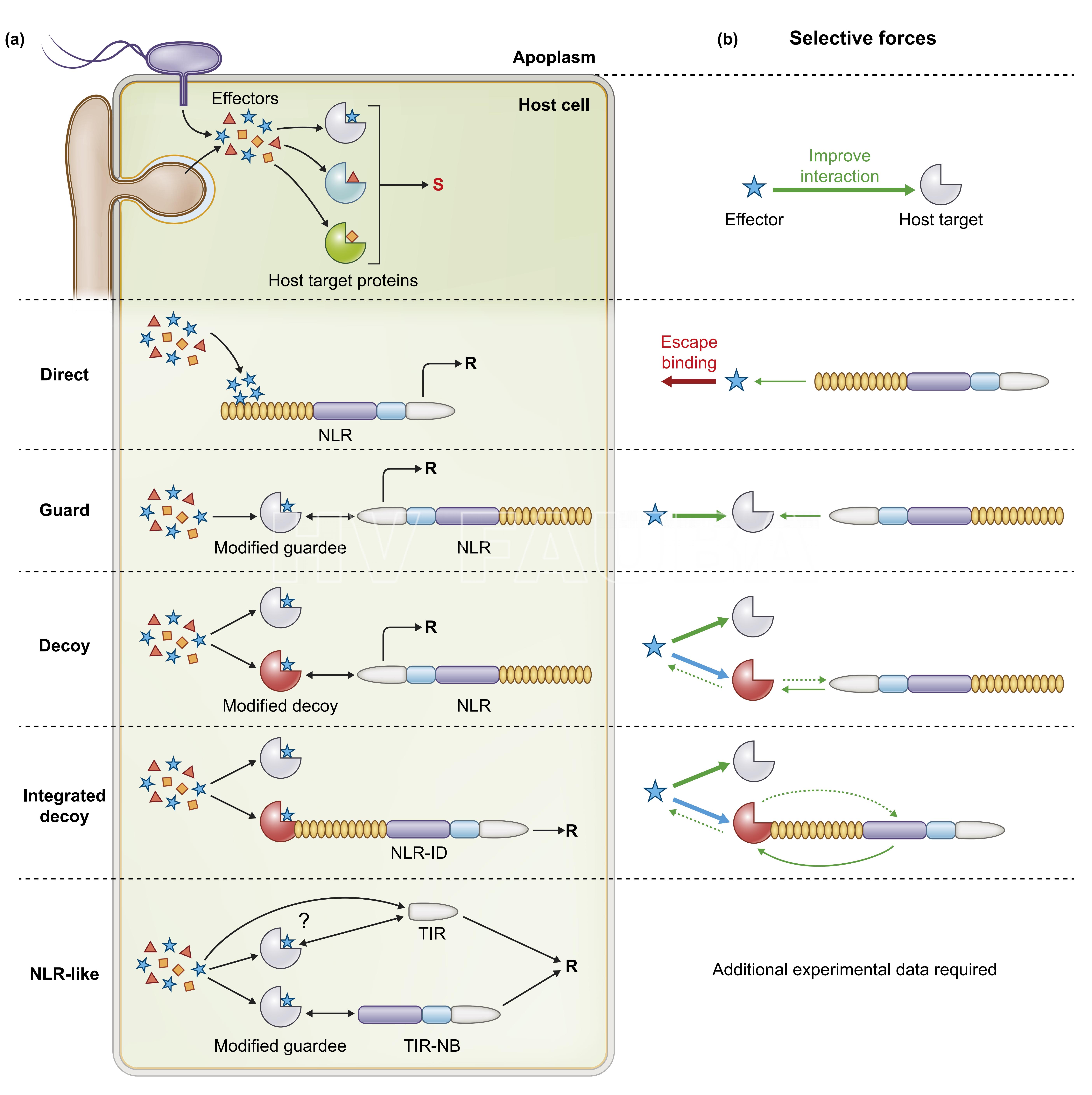
- Fig. 3 . Representación del funcionamiento de la interacción efector-receptor immune NLR (Cesari, 2018).
Figura 3. Representación conceptual esquemática de los probables funcionamientos de la interacción efector–receptor immune (Cesari, 2018). Los receptores inmunes NLR detectan efectores patógenos específicos a través de diversos mecanismos con diferentes restricciones evolutivas.
(a) Tras la infección, los efectores patógenos se dirigen y modifican las proteínas del hospedante para promover la susceptibilidad (S). La interacción directa NLR–efector induce resistencia (R). El reconocimiento indirecto se produce cuando las proteínas efectoras target (objetivo) están protegidas por los NLR (la proteína target u objetivo se llama guardee o de vigilancia) o si los genes target duplicados evolucionan en proteínas señuelo monitorizadas por los NLR del hospedante. En estos modelos indirectos, los NLR detectan modificaciones mediadas por efectores de proteínas guardee o señuelo o de vigilancia. Los señuelos o proteínas de vigilancia (guardee) a menudo se integran en la estructura de los NLR, lo que permite la detección de efectores mediante el enlace directo. Algunas proteínas similares a NLR, que carecen de dominios reguladores como el NB y el LRR o el LRR únicamente, también funcionan como sensores de efectores pero su modo de acción no está aún del todo clarificado.
(b) Los efectores evolucionan hacia la interacción con los target del hospedante (incluidas las proteínas guardee o de vigilancia) mientras que los target no muestran evidencia para la selección positiva, probablemente porque la variación puede perjudicar su función central en la inmunidad o fisiología de las plantas. En el modelo directo de reconocimiento de efectores, los efectores mutan para escapar de la unión de NLR mientras que los NLR están bajo presión selectiva para mantener o restaurar esta unión. Los modelos de guardia, señuelo y señuelo integrado pueden ofrecer opciones de detección de efectores más efectivas para el sistema inmune de la planta que el modelo directo. De hecho, a través de su función de virulencia, los efectores pueden verse obligados a evolucionar hacia el reconocimiento y pueden no evadir directa o fácilmente la detección sin modificar su actividad. Las proteínas señuelo no tienen ninguna función además de la detección efectora y, por lo tanto, podrían evolucionar hacia unión efectora y NLR. En el modelo de señuelo integrado, la proximidad del NLR al objetivo efector se mantiene mediante una conexión física, pero el ID y otros dominios de la proteína NLR aún pueden coevolucionar para mantener la interoperabilidad. Flechas gruesas = evolución efectora; flechas discontinuas = señuelo o evolución de señuelo integrado; flechas delgadas = evolución NLR; flechas verdes = fuerzas de selección que mejoran la interacción; flechas rojas = fuerzas de selección que disminuyen la interacción; flechas azules = evolución indirecta de efectores hacia señuelo o interacción de señuelo integrada.
.
La homeostasis apropiada de la actividad de los NLR es crítica para su función (Li et al., 2015). Los niveles de proteína NLR deben estar por encima de un cierto umbral para activar suficientemente la señalización de defensa. Sin embargo, las mutaciones que activan de manera constitutiva los receptores inmunes NLR o dan como resultado niveles elevados de expresión de NLR pueden conducir a la letalidad embrionaria, la muerte celular espontánea y / o el retraso en el crecimiento de las plantas (Lai & Eulgem, 2017). La resistencia mediada por NLR puede asociarse con una reducción sustancial de la aptitud física de la planta (Tian et al., 2003).
.
Efectores patogénicos
Los efectores, tales como las toxinas y las proteínas efectoras, se pueden definir como moléculas derivadas de patógenos destinadas a promover la virulencia de los patógenos mediante la interacción con el hospedante. En otras palabras, los efectores son factores de virulencia que generalmente no tienen una función de gen constitutivo de crecimiento y desarrollo fuera del hospedante. Por supuesto, cuando un efector es reconocido por un receptor de defensa del hospedante, la función de virulencia deseada a menudo se ve ensombrecida por una función de avirulencia dominante (Bent & Mackey. 2007).
Las moléculas efectoras de patógenos se translocan en células y tejidos del hospedante, donde se dirigen a macromoléculas importantes (proteínas, componentes de la pared celular, ácidos nucleicos, etc.) involucradas en funciones celulares normales y/o relacionadas con la inmunidad (De Wit et al., 2009). Los efectores generalmente se catalogan en dos grupos distintos: los que actúan en los espacios extracelulares de los tejidos del hospedante (espacio apoplástico) o los que actúan dentro de las células del hospedante (espacio citoplasmático). Los efectores apoplásticos se secretan a través de sistemas de secreción eucarióticos generales en oomicetes y hongos (Manning & Ciuffetti, 2005), o mediante haustorios (Voegele & Mendgen, 2003; Garnica et al., 2014), o mediante el sistema de secreción de tipo II (T2SS) o utilizando un sistema especializado de secreción tipo III (T3SS) de patógenos bacterianos (Henry et al., 2017). Estas moléculas típicamente están involucradas en la degradación enzimática de las paredes celulares de las plantas, la evasión inmune o la supresión de la actividad proteolítica del hospedante (Carella et al., 2018).
Después de la translocación al interior de las células vegetales, algunos efectores microbianos pueden dirigirse a diferentes compartimentos subcelulares. Los efectores citoplasmáticos han sido ampliamente estudiados en angiospermas, revelando un conjunto de estrategias de virulencia donde los efectores acceden a varios compartimentos celulares (núcleo, cloroplasto, citoplasma, etc.) para interrumpir la actividad de las proteínas del hospedante, involucradas en la regulación transcripcional, secreción, metabolismo, muerte celular programada y señalización hormonal (Carella et al., 2018). Asimismo, curiosamente, un número significativo de proteínas efectoras de diferentes microorganismos patógenos, incluyendo virus, oomycetes, hongos, nemátodos y bacterias, se dirigen al núcleo de las células hospedantes, donde efectúan su acción de virulencia (Rivas & Genin, 2011).
Los genes que codifican dichos efectores se encuentran entre los genes que evolucionan más rápidamente en genomas de patógenos (Dodds, 2010). Análisis de genómica poblacional mostraron que los patógenos que evolucionan rápidamente poseen altas tasas de recambio en los loci efectores y muestran un mosaico en la presencia / ausencia de polimorfismo en los efectores entre las diversas cepas. Las estrategias eficaces de patógenos requieren un potencial del patógeno para la adaptación rápida (Sanchez-Vallet et al., 2018).
.
Efectores patogénicos de hongos
Los patógenos tienen éxito a través del despliegue espaciotemporal de una gama notablemente diversa de proteínas efectoras para controlar las defensas de las plantas y los procesos celulares (de Jonge et al., 2011). Algunos efectores pueden ser secretados por apresorios incluso antes de la penetración del hospedante, muchos ingresan a las células vegetales vivas donde se dirigen a diversos compartimentos subcelulares y otros se mueven hacia las células vecinas para prepararlos antes de la invasión (Giraldo & Valent, 2013).
La mayoría de los hongos biotróficos tienen la capacidad de formar haustorios, una estructura especializada en la infección (Voegele & Mendgen, 2003). Una vez dentro del tejido vegetal, la invasión celular se logra mediante diversos tipos de hifas biotróficas. Algunos patógenos crecen de una célula vegetal a la siguiente como hifas invasivas intracelulares (IH), algunos crecen solo como hifas extracelulares y otros crecen como hifas extracelulares que insertan haustorios, que son estructuras de alimentación terminales, dentro de las células del hospedante. Contrariamente a las bacterias Gram negativas, no se ha descubierto ningún TTSS para hongos patógenos, y sus efectores se pueden liberar a partir de los haustorios en el espacio intercelular (apoplasto) de la planta. Los haustorios no solo son estructuras especializadas de alimentación necesarias para la adquisición de nutrientes, sino que también inducen cambios estructurales, celulares y bioquímicos en las células hospedantes invadidas (Garnica et al., 2014). Además, pueden facilitar la entrega de efectores en la matriz extrahaustorial, varios de los cuales se trasladan posteriormente a las células hospedantes (Stergiopoulos & de Wit, 2009).
El determinante Avr4 contiene un dominio obligatorio que une quitina, el componente mayoritario de las paredes celulares de los hongos. Como mecanismo para percibir la quitina como PAMP, las plantas desarrollaron probablemente quitinasas para aumentar los polímeros activos de las membranas celulares de patógenos invasores, de manera tal que accionaran las respuestas de defensa. Para evadir el reconocimiento y la activación de las respuestas específicas de las plantas inducidas por quitina, el Avr4 de C. fulvum se une a las quitinasas del hongo en la pared celular (Jones et al., 1994; Thomas et al., 1997).
En el caso de los pseudohongos, el análisis del genoma de algunas especies reveló la presencia de cientos de efectores diferentes en especies individuales de Phytophthora (Tyler et al., 2006; Haas et al., 2009).
.
Efectores patogénicos de bacterias
Se conoce que los patógenos bacterianos de animales pueden secretar solamente un número limitado de efectores dentro de las células del hospedante. Sin embargo, los patógenos de las plantas, tales como la bacteria Pseudomonas syringae, pueden secretar aproximadamente de 20 a 30 efectores durante la infección. Los efectores promueven la patogenicidad, y el TTSS es esencial para el desarrollo de los síntomas de la enfermedad y la multiplicación bacteriana. Por su acción colectiva, se ha considerado la hipótesis de que los efectores alteran la fisiología vegetal en hospedantes susceptibles para sostener el crecimiento del patógeno.
Las proteínas efectoras de hongos y bacterias, que se liberan a las plantas pueden poseer actividad enzimática (Tabla 1). Estas enzimas son responsables de modificar las proteínas del hospedante para aumentar la virulencia del patógeno y evadir su detección. Los patógenos se deben proteger contra estas actividades enzimáticas efectoras potencialmente perjudiciales.
Del gran número de proteínas efectoras bacterianas que han sido clonadas, sólo unas pocas se han caracterizado bioquímicamente.
.
Efectores patogénicos de virus
El principal medio a través del cual las plantas se defienden contra la infección viral es el silenciamiento del ARN viral, que regula la acumulación de moléculas endógenas y exógenas. La mayoría de los virus de las plantas tiene genomas de ARN, y las estructuras secundarias de doble cadena pueden accionar el silenciamiento del genoma viral en el hospedante, y prevenir la diseminación viral sistémica. Los efectores de la virulencia viral se pueden considerar los encargados de suprimir la respuesta silenciadora del ARN del hospedante.
Se han identificado supresores en muchos virus de las plantas, y las funciones moleculares de varios de ellos han sido caracterizadas detalladamente. Estos estudios determinaron que diversos supresores interfieren con los componentes únicos de la maquinaria silenciadora del hospedante, lo que sugiere que muchos virus desarrollaron independiente los medios para suprimir el silenciamiento (Li & Wang, 2018). Naturalmente, las plantas utilizan un segundo mecanismo de defensa para reconocer y para restringir el movimiento del virus.
Las proteínas R específicas reconocen componentes virales de cualquier supresor y lo silencian; además pueden reconocer otras proteínas que se acumulen como resultado de la replicación viral.
.
En resumen, aunque diversas bacterias, hongos y virus patógenos de plantas codifican diferentes proteínas Avr, poseen diferentes elicitores o inductores y usan distintas estrategias de infección, parecen alterar de manera similar los estados metabólicos, fisiológicos y celulares de los hospedantes. Además, los componentes de señalización como los complejos SGT1 / RAR1 / HSP90 y EDS1 / PAD4 / SAG101, sirven de manera similar para mediar las respuestas de resistencia contra diversos patógenos virales y no virales, lo que sugiere que las plantas han desarrollado resistencia a enfermedades de amplio espectro para defenderse simultáneamente contra diversos tipos de patógenos (Mandadi & Scholthof, 2013).
.
Tabla 1. Ejemplos de actividad enzimática de efectores y activadores de hongos y bacterias liberados contra las plantas (Chisholm et al., 2006 modificado).

Hormonas
Las hormonas vegetales desempeñan un papel importante en la regulación de los procesos de desarrollo y las redes de señalización involucradas en las respuestas de las plantas a una amplia gama de estreses bióticos y abióticos (Bari & Jones, 2009; Pieterse et al., 2009; Gasperini y Howe, 2024).
Muchos patógenos producen pequeñas moléculas efectoras que mimetizan las hormonas de las plantas. Existen tres moléculas señalizadoras que regulan la defensa de la planta contra los ataques microbianos: el ácido salicílico (SA), el ácido jasmónico (JA), y el etileno (Et). La señalización Et-dependiente es importante para la respuesta de la planta ante el ataque de los patógenos, ante una herida mecánica y/o inducida por herbívoros. La señalización SA-dependiente es crítica en el establecimiento de resistencia bacteriana local y sistémica, mientras que la señalización JA-dependiente se induce en respuesta a la herida y a la depredación mecánica de herbívoros. Las rutas de señalización de la defensa a través del SA y de JA son, generalmente, antagónicos, y los patógenos bacterianos se han desarrollado para suprimir las respuestas de la defensa mediadas por el SA.
El sistema de defensa de la planta se afina y se automodula cuidadosamente para responder a los diferentes comportamientos de infección de los diversos patógenos microbianos (Andolfo & Ercolano, 2015).
.
.
Reconocimiento directo e indirecto de efectores patogénicos
Resistencia gen-por-gen
Como se ha descrito anteriormente, la evolución de las proteínas efectoras secretadas por los patógenos de plantas llevó en última instancia a la adquisición de proteínas que reconocen específicamente estos determinantes bacterianos, fúngicos y virales. Esta asociación en parejas que describe el reconocimiento de efectores dentro de la célula de la planta se ha caracterizado genéticamente como resistencia gen-por-gen. En presencia de una asociación efector-proteína R, la resistencia se activa, dando como resultado la iniciación de la señalización de defensa y una respuesta de defensa o resistencia del hospedante. La resistencia se manifiesta como muerte celular localizada en el sitio de la infección, conocida como reacción de hipersensibilidad (HR) y la inhibición del crecimiento del patógeno. Inversamente, en la ausencia de esta interacción, el patógeno elude la detección por la planta, dando por resultado la infección, la proliferación del patógeno y el inicio de la enfermedad.
También se ha encontrado evidencia de que, en lugar de desarrollar receptores para cada efector posible, las plantas hospedantes han desarrollado mecanismos para monitorear los objetivos comunes del hospedante. Al monitorear las perturbaciones, las proteínas R detectan indirectamente la actividad enzimática de múltiples efectores (Van der Biezen & Jones, 1998; Chisholm et al., 2006).
.
Resistencia por arquitectura de dominios proteicos
Los efectores introducidos por los patógenos que evaden la PTI son reconocidos por los genes específicos R, la mayoría de los cuales codifican para las proteínas NB-LRR (del Inglés Nucleotide-binding/leucine-rich repeat (NLR) immune receptors = receptores inmunitarios de unión a nucleótidos / repetición rica en leucina) que constituyen la clase mayoritaria de proteínas resistentes de la resistencia gen-por-gen de las plantas. Más de 30 genes codifican para este tipo de proteínas. Esta familia multigénica ha sido aislada en varias especies de plantas, y constituyen un mecanismo de defensa eficiente que provee resistencia frente a diversos patógenos como insectos, nemátodos, hongos, pseudohongos, bacterias, virus y viroides. Estas proteínas de resistencia se expresan constitutivamente en la mayoría de las células con potencialidad de ser atacadas por patógenos, de esta manera se compensa la ausencia de un sistema circulatorio en las plantas.
Numerosas proteínas R han sido identificadas. Son moléculas citoplasmáticas, y además de las regiones NB y LRR mencionadas con anterioridad tiene varios dominios efectores en el extremo N-terminal. Análisis comparativos de proteínas NB-LRR mostraron que los residuos de lámina â polares expuesto son hipervariables y sujetos a una presión selectiva para su diversificación, lo que indica que la especificidad de reconocimiento reside en esta parte de los LRR.
La región N-terminal está involucrada en la cascada de señalización, mientras que los sitios LRR, parecen estar implicados en la formación de interacciones proteína-proteína y en el reconocimiento específico del efector.
Las proteínas NB-LRR pueden servir como moléculas adaptadoras que unen el reconocimiento con la transducción de la señal. Por ejemplo, cuando las señales son percibidas por los LRR, se inicia la hidrólisis nucleotídica del dominio NB. Esto puede proveer la energía necesaria para un cambio conformacional en la proteína NB-LRR, y se expone la porción efectora N-terminal, lo que dispararía la respuesta de defensa. La activación de NB-LRR produce una red de interrelaciones entre las vías de respuestas inmunes activadas, en parte, para diferenciar el ataque de patógenos biotróficos y necrotróficos. Esto se mantiene por el equilibrio entre el SA, una señal local y sistémica para la resistencia contra muchos biótrofos, y la acumulación de la combinación de JA y del etileno como señales que promueven la defensa contra necrotrofos. La activación de NB-LRR induce respuestas diferenciales entre el SA y respuestas dependientes de especies reactivas del oxígeno. El comienzo de las oxidaciones de oxidasa NADPH-dependiente que acompaña a la activación de la ETI reprime la muerte celular SA-dependiente en las células circundantes al sitio de la infección. Los cambios locales y sistémicos en la expresión genética son mediados, en gran medida, por los factores de transcripción de la familia WRKY y TGA. La reprogramación transcriptiva de las células de la planta después de un ataque patógeno es extensa, y afecta entre el 3 y 12% de los 24.000 genes evaluados de A. thaliana luego de ataques de hongos y bacterias, respectivamente (Thilmony et al., 2006; Wang et al., 2011; Tsuda y Somssich, 2015; Li et al., 2016; Bjornson et al., 2021; Thieffry et al., 2022; Zhang et al., 2022).
Varias proteínas NB-LRR reconocen efectores tipo III indirectamente, a través de la detección de los productos de su acción en blancos del hospedante, consistentemente con la ‘hipótesis guardiana’. Los principios dominantes de esta hipótesis son: (1) un efector actúa como factor de virulencia cuando tiene un blanco en el hospedante; (2) el efector contribuye al éxito del patógeno y altera este blanco en genotipos susceptibles del hospedante; y (3) la perturbación del blanco en el hospedante, genera una modificación propia patógeno-inducida del patrón molecular, que activa las proteínas NB-LRR correspondiente. De este modelo se derivan consecuencias importantes: múltiples efectores podrían desarrollarse de forma independiente para manipular el mismo blanco en el hospedante, lo cual podría conducir a la evolución de más de una proteína NB-LRR asociada a un blanco de efectores múltiples; además, estas proteínas NB-LRR serían activadas por el reconocimiento de diversos patrones propios modificados producidos en el mismo blanco por la acción de diversos efectores.
Una segunda clase importante de genes R codifica para las proteínas extracelulares LRR (eLRR). Tres subclases de eLRR se han clasificado según sus estructuras de dominio. Estas subclases incluyen proteínas RLP (del inglés: receptor-like proteins) con un domino LRR extracelular y uno transmembrana (TM) (Kruijt et al., 2005), proteínas RLK (del inglés: receptor-like kinases) con un dominio TM y una quinasa citoplasmática (Shiu & Bleecker, 2001a; Gish et al., 2011; Liu et al., 2017) y la proteína PGIP (del inglés: polygalacturonase inhibiting protein) con un dominio LRR en la pared celular (De Lorenzo et al., 2001; Di Matteo et al., 2006).
.
Proteínas similares a los receptores (RLP) involucradas en respuestas desencadenadas por efectores fúngicos apoplásticos
Las proteínas similares a los receptores (RLPs, del inglés: Receptor-like proteins) comparten similitud estructural con las quinasas tipo receptor (RLK, del inglés: receptor-like kinase) pero carecen de un dominio citoplásmico de quinasa (Stotz et al., 2014; Jamieson et al., 2018). Se han identificado RLK con diversos dominios extracelulares de reconocimiento de ligandos (por ejemplo, repeticiones ricas en leucina, tipo lectina/lectina, tipo factor de crecimiento epidérmico, etc.). Las RLK que contienen repeticiones ricas en leucina (LRR-RLK) son las RLK más comunes y las más estudiadas en las plantas. De forma similar, el dominio de unión a ligando extracelular más común que se encontró en las RLP es LRR (Shiu y Bleecker, 2003). Existen aproximadamente 223 LRR-RLK y 57 LRR-RLP en Arabidopsis (Shiu y Bleecker, 2001b; Fritz-Laylin et al., 2005; Wang et al., 2008; Lehti-Shiu et al., 2009). En las LRR-RLK, el dominio de quinasa intracelular exhibe más conservación que el dominio de LRR extracelular.
.
-
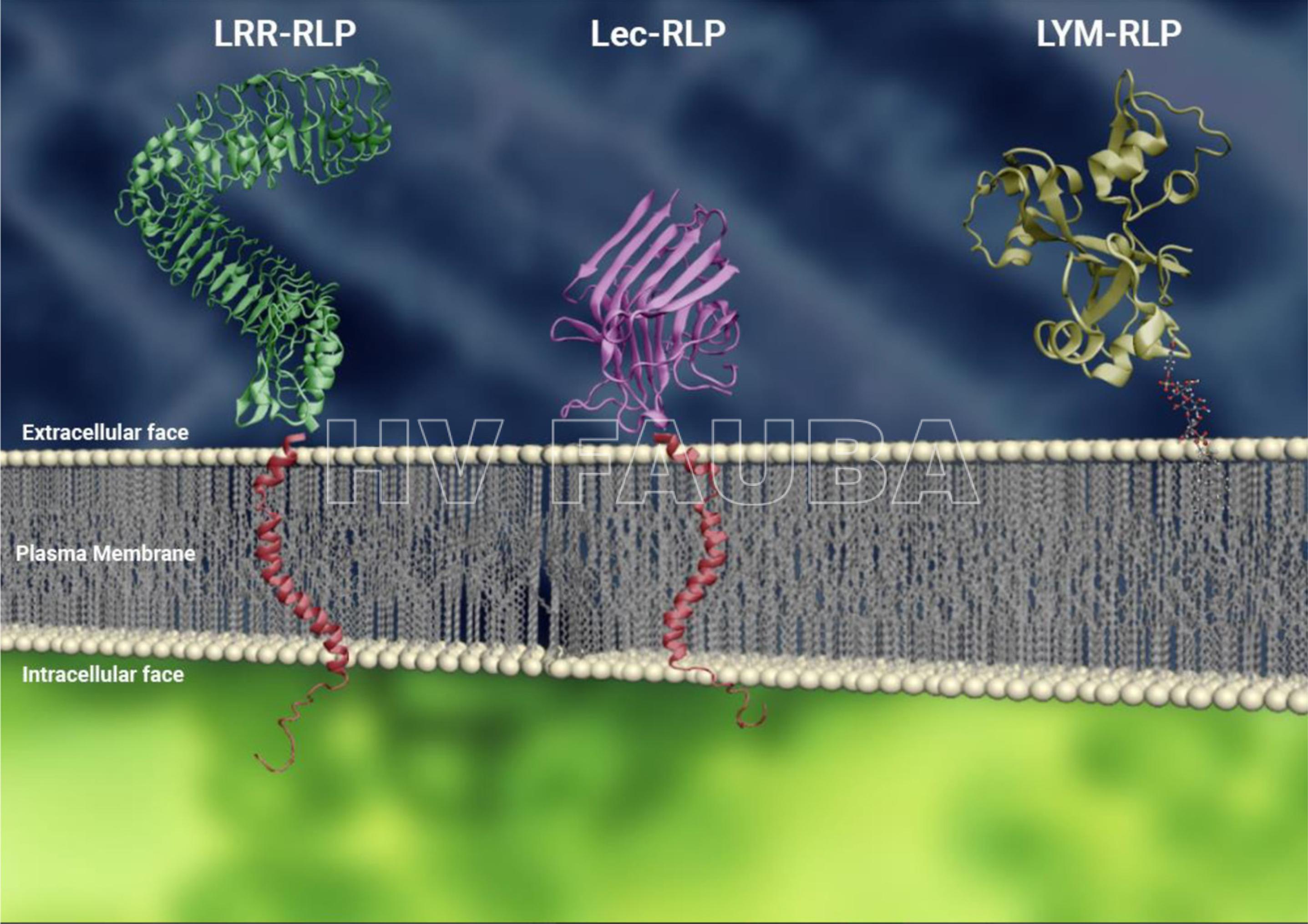
- Fig. 4. Representaciones conceptuales de RLPs con diferentes ectodominios en la membrana plasmática.
Figura 4. Representaciones conceptuales de RLPs con diferentes ectodominios en la membrana plasmática. De izquierda a derecha: LRR-RLP (verde), Lec-RLP (púrpura) y LYM-RLP (dorado); hélices transmembrana se representan en rojo. Las hélices transmembrana se añadieron para representar la estructura, pero no representan la estructura real de los dominios transmembrana de estas proteínas. El área intracelular se indica adicionalmente mediante la representación de colas cortas citoplásmicas (rojas) (Jamieson et al., 2018).
.
La primer RLP que fue caracterizada en tomate, Cf-9, codifica para una LRR-RLP (Jones et al., 1994). Cf-9 confiere resistencia específica a las razas de Cladosporium fulvum que llevan el gen efector Avr9. Esta relación se caracterizó como una resistencia de tipo «gen por gen», que ahora se denomina ETI. Luego, se demostró que una segunda LRR-RLP de tomate designada Cf-4 era necesaria para la resistencia a las razas de C. fulvum que expresan el gen efector Avr4 (Thomas et al., 1997). Mientras que la percepción de MAMP/PAMP por RLK o RLP de acuerdo con el modelo vigente aceptado solo conduce a la activación de la PTI, el reconocimiento de Avr4 y Avr9 dá como resultado la activación de HR, una respuesta que es más comúnmente asociada con la ETI (Jones et al., 1994; Thomas et al., 1997). Además, Cf-9 no parece percibir directamente el Avr9, ya que no existe una unión directa entre ellos (Luderer et al., 2001). Por lo tanto, como indica su nomenclatura (Avr para «avirulencia»), se clasifica a Avr4 / Avr9 como genes que codifican para efectores apoplásticos, en lugar de MAMP/PAMP (Stotz et al., 2014). Este ejemplo confunde o no encaja de manera perfecta en el esquema tradicional de interacción R-gen / Avr-gen, ya que Cf-4 y Cf-9 son estructuralmente similares a los PRR, pero se definen funcionalmente como genes R. En consecuencia, se considera que el reconocimiento de efector apoplástico por RLP activa la «defensa desencadenada por efector» (ETD, del inglés: effector-triggered defense). ETD se distingue de ETI no solo por la ubicación del reconocimiento de los efectores, sino también por su mayor tiempo de respuesta, propensión a la activación mediada por heterodímeros y la tasa de muerte celular (Stotz et al., 2014).
.
.
El modelo de zig-zag de Jones y Dangl (2006) se ajusta bien para ilustrar la producción cuantitativa del sistema inmunológico de las plantas, funcionando como un modelo expositivo de la historia evolutiva del sistema inmunológico de las plantas. Sin embargo, el modelo tiene limitaciones que significan que no es un marco cuantitativo o predictivo para el estudio directo de las interacciones planta-microbio (Pritchard y Birch, 2014; Liu et al., 2021).
.
.
Resistosomas
Los receptores inmunitarios de unión a nucleótidos / repetición rica en leucina (NLR) detectan los efectores de patógenos y desencadenan la respuesta inmunitaria de una planta. Los NLR reconocen la presencia o actividad celular de proteínas de virulencia derivadas de patógenos, los denominados efectores. La activación de NLR dependiente de efector conduce a la formación de complejos oligoméricos de orden superior, denominados resistosomas (Burdett et al., 2019; El Kasmi, 2021). Los resistosomas pueden formar canales de cationes potenciales permeables al calcio en las membranas celulares e iniciar afluencias de calcio que dan como resultado la activación de la inmunidad y la muerte celular o funcionar como NADasas cuya actividad es necesaria para la activación de componentes de señalización inmunes aguas abajo, dependiendo del dominio N-terminal de la proteína NLR.
Recientemente se han definido las estructuras de dos NLR que portan dominios del receptor de interleucina 1 (TIR) de tipo Toll (TIR-NLR) (Li et al., 2020; Lian et al., 2021) y de la proteína NLR de cola enrollada en espiral (CC) – RESISTENCIA ACTIVADA POR HOPZ 1 (ZAR1) formando un estado oligomérico (Wang et al., 2019a, 2019b). Ma et al. (2020) estudiaron el TIR-NLR RPP1 en Arabidopsis thaliana que reconoce Peronospora parasitica 1 y su respuesta a los efectores de este oomicete patógeno. Martin et al. (2020) estudiaron el TIR-NLR ROQ1 en Nicotiana benthamiana que reconoce XopQ 1 y su respuesta al efector de Xanthomonas. Estos estudios encontraron que estos TIR-NLR formaban tetrámeros que, cuando se activan al unirse al efector del patógeno, exponen el sitio activo de una nicotinamida adenina dinucleósido (NAD) hidrolasa. Por tanto, el reconocimiento del efector patógeno inicia la hidrólisis de NAD y comienza la respuesta inmune. La señalización NLR en plantas, como lo ilustra la estructura ZAR1, es análoga a los mecanismos de señalización del receptor inmunitario innato en todos los reinos (Maekawa et al., 2011; Jones et al., 2016; Burdett et al., 2019; Steele et al., 2019).
Las vías inmunes activadas por los receptores de la superficie celular e intracelulares en las plantas se potencian mutuamente para activar defensas fuertes contra los patógenos (Ngou et al., 2021). Estos hallazgos modifican nuestra comprensión de la inmunidad de las plantas y tienen amplias implicaciones para la mejora de los cultivos.
.
.
Activadores endógenos
Los activadores endógenos en plantas son típicamente fragmentos de oligosacáridos, tales como los derivados de membranas celulares de la planta por las carbohidrasas producidas por los patógenos o por las enzimas endógenas de la planta, que se sintetizan en respuesta a ataques de patógenos.
.
Evasión de la ETI por parte de los patógenos
La ETI es menos efectiva para los microorganismos que pueden evadir el reconocimiento mediado por las NB-LRR de un efector particular. Las frecuencias alélicas del efector son probablemente influenciadas por su modo de acción. Los alelos de estos efectores tienen un alto nivel de selección diversificada, probablemente, son seleccionados por el reconocimiento del hospedante, y por lo tanto actúan en los residuos efectores que no se requieren para la función efectora.
La activación de las proteínas NB-LRR por la vía del reconocimiento patógeno-inducido proporciona modificaciones propias, y facilita un mecanismo para el reconocimiento de múltiples efectores involucrados en comprometer el mismo blanco del hospedante. Para que la selección genere un efector que escape a la ETI, el efector tiene que tener la probabilidad de perder su función nominal. La respuesta más simple del patógeno al reconocimiento del hospedante es desechar el gen efector detectado, con tal de que el repertorio de la población efectora pueda cubrir la pérdida potencial de aptitud en los hospedantes susceptibles. De hecho, los genes efectores se asocian a menudo a los elementos genéticos móviles o a los telómeros y se observan comúnmente como presencia/ ausencia de polimorfismo a través de cepas bacterianas y fúngicas.
.
La ETI se puede superar también con la evolución de los efectores del patógeno que la suprimen directamente.
La evolución microbiana en respuesta a la ETI puede dar lugar a dos extremos de la evolución de NB-LRR. Algunos genes de NB-LRR no son duplicados y se han desarrollado de manera relativamente lenta, sus productos quizás se asocian establemente con proteínas del hospedante y retardan la diversificación. Otros microorganismos se están desarrollando más rápido y pueden interactuar directamente con efectores en plena evolución. En poblaciones de patógenos, la frecuencia de un gen efector podrá ser aumentada según su capacidad de promover virulencia, y reducida por el reconocimiento del hospedante. Por lo tanto, la selección natural debe mantener la función efectora en ausencia del reconocimiento. Pero la función efectora tiene un costo dependiente de la frecuencia de genes R correspondientes. Así, si la frecuencia efectora cae en una población del patógeno, las plantas hospedantes se pudieran seleccionar para la pérdida del alelo R correspondiente, y el ciclo dependiente de la frecuencia continuaría.
.
Transcripción de Genes relacionados con la respuesta inmune
La inmunidad de las plantas depende de la expresión masiva de los genes relacionados con la patogénesis (PR, del inglés: pathogenesis-related genes), cuya transcripción es desreprimida por señales inducidas por patógenos. La hormona vegetal ácido salicílico (SA) actúa como una molécula de señalización importante en la inmunidad de las plantas y en la resistencia sistémica adquirida provocada por patógenos. La señal de SA da como resultado la activación del regulador inmunitario «maestro», denominado: Nonexpressor of pathogenesis-related genes 1 (NPR1), que se cree que es reclutado por factores de transcripción como los TGA (TGACG-BINDING FACTORs) a numerosos PR en sentido descendente. A pesar de su papel clave en la inmunidad activada por SA, la naturaleza bioquímica de la función coactivadora transcripcional de NPR1 y la reprogramación masiva de la transcripción inducida por ella todavía no está del todo esclarecida. Hasta el momento se ha demostrado que los coactivadores transcripcionales que controlan una variedad de procesos de diferenciación y desarrollo como los HATs (del inglés: CBP/p300-family histone acetyltransferases) y NPR1 forman un complejo coactivador y se reclutan para la cromatina de PR a través de TGA en la señal de SA, y finalmente el complejo HAC-NPR1-TGA activa la transcripción de genes PR mediante la reprogramación epigenética mediada por acetilación de histonas. Este mecanismo revela un funcionamiento molecular de la reprogramación transcripcional mediada por NPR1 y un aspecto epigenético clave del sistema inmunitario central en las plantas, donde los HACs y NPR1 son esenciales para desarrollar la respuesta inmunidad activada por SA y la inducción de genes PR (Jin et al., 2018).
.
Reacción de Hipersensibilidad, Autofagia y Muerte celular programada
La autofagia es un proceso catabólico altamente conservado en eucariotas, en el cual el citoplasma, incluyendo el exceso de orgánulos o aquellos deteriorados o aberrantes, son secuestrados en vesículas de doble membrana y liberados dentro del lisosoma/vacuola para su descomposición y eventual reciclado de las macromoléculas resultantes. En las plantas, el proceso catabólico altamente conservado de la autofagia se conoce desde hace tiempo como un medio para mantener la homeostasis celular y hacer frente a las condiciones de estrés abiótico. La acumulación de pruebas ha vinculado la autofagia a la inmunidad contra los patógenos invasores, la regulación de la muerte celular de las plantas y las defensas antimicrobianas. A su vez, parece que los fitopatógenos han evolucionado en formas no solo de evadir el aclaramiento autofágico sino también de modular y manipular el proceson de autofagia vegetal para su propio beneficio (Leary et al., 2018).
El reconocimiento de patógenos a menudo desencadena una respuesta de hipersensibilidad (HR, del inglés: hypersensitive response), la liberación rápida de compuestos antimicrobianos y muerte celular programada (PCD, del inglés: programmed cell death) local. La HR suprime de manera eficiente el crecimiento de la mayoría de los patógenos biotróficos. Algunos inmunorreceptores de superficie especializados y un conjunto de inmunorreceptores citoplásmicos / intracelulares conocidos como proteínas con dominio de unión a nucleótidos y que contienen repeticiones ricas en leucina (NLRs, del inglés: nucleotide-binding domain and leucinerich repeat-containing proteins) pueden detectar las proteínas efectoras secretadas por patógenos (Jacob et al., 2013). La activación de las NLRs inicia la inmunidad desencadenada por efectores (ETI) que a menudo va acompañada de una respuesta celular de hipersensibilidad (HR, del inglés: hypersensitive response) (Duxbury et al., 2016; Wu et al., 2017). El reconocimiento de efectores por NLR es principalmente indirecto y con frecuencia implica la modulación de las proteínas del hospedante dirigidas por efectores protegidos por los NLR. Por lo tanto, el despliegue preciso de los receptores inmunes, sus “guardianes”, y componentes de defensa en sitios particulares y en cantidades correctas es crítico no solo para el reconocimiento inmunitario sino también para la ejecución de mecanismos de defensa que conducen a la eliminación de patógenos.
La HR suprime de manera eficiente el crecimiento de la mayoría de los patógenos biotróficos pero al mismo tiempo facilita la colonización de la planta por patógenos necrotróficos, como por ejemplo B. cinerea y S. sclerotiorum (Mbengue et al., 2016). Para algunos patógenos necrotróficos como por ejemplo B. cinerea y S. sclerotiorum, se propuso que la secreción de ácido oxálico y proteínas que provocan la muerte celular promueven la virulencia mediante la inducción de PCD del hospedante. Los mecanismos moleculares por los que B. cinerea y S. sclerotiorum manipulan la PCD del hospedante permanecen aún no del todo elucidados. Además, los mutantes de Arabidopsis alterados en la autofagia, una forma de PCD que implica la degradación de componentes citoplásmicos, fueron más susceptibles a B. cinerea (Lai et al., 2011). Además, los mutantes deficientes en ácido oxálico de S. sclerotiorum desencadenan la muerte celular autofágica restringida, lo que sugiere que el ácido oxálico puede suprimir la fisiopatología en las células hospedantes (Kabbage et al., 2013). Estos hallazgos indican que ciertos mecanismos de muerte de la célula hospedante pueden ser favorable a los hongos necrotróficos mientras que otros serían perjudiciales (Dickman y de Figueiredo, 2013).
El control de la PCD en las células fúngicas también desempeña un papel crucial en la patogenicidad. Se observó que la PCD fúngica es atenuante en las etapas iniciales de la infección por B. cinerea, y se encontró que la proteína anti-apoptótica BcBIR1 contribuye positivamente a la virulencia de B. cinerea. Estos hallazgos sugieren que la PCD fúngica podría ser desencadenada por moléculas de defensa de las plantas, y que se requiere maquinaria fungicida anti-apoptótica para prevenirla (Shlezinger et al., 2011). Esto coloca el control de los programas de muerte celular programada de plantas y patógenos fúngicos en el centro de la carrera durante la interacción de las plantas con los hongos necrotróficos.
.
Resistencia sistémica adquirida
Una primera inoculación con un patógeno que permanece localizado en el sitio de infección puede conducir a la resistencia contra una infección posterior. Esta resistencia se expresa localmente en el sitio de infección inicial, pero también sistémicamente, en tejidos intactos (Conrath et al., 2006; Jung et al., 2009; Zhang et al., 2021). Por ejemplo, la infección de una hoja conduce a la protección en las partes no infectadas de la misma hoja pero también a la resistencia sistémica en otras partes de la planta. Los tejidos de las plantas que exhiben resistencia sistémica adquirida (SAR, del inglés: systemic acquired resistance) en las partes distantes libres de patógenos muestran un estado «preparado» (cebado o activado) de defensa asociado con mecanismos de defensa más rápidos y más fuertes. La respuesta SAR se genera por una señal en hojas infectadas primarias y el movimiento de la señal hacia órganos distantes (Schneider et al., 1996; Grant & Lamb, 2006; Klessig et al., 2018).
La secuencia de eventos que permiten la respuesta sistémica comienza localmente. En las células adyacentes a la respuesta hipersensitiva se observa el engrosamiento de las paredes celulares por incorporación de proteínas estructurales o lignina, deposición de calosa y la inducción de la síntesis de fitoalexinas. En las células más distantes, o sea las partes no infectadas de la planta, las primeras reacciones de defensa tipo SAR incluyen la síntesis de proteínas relacionadas con la patogénesis (PR), enzimas (β-1,3 glucanasas, endohidrolasas, quitinasas), inhibidores de enzimas como la taumantina, inhibidores de amilasa y proteinasas, entre otras. Los genes que son inducidos en las infecciones primarias por el patógeno, se expresan localmente y también sistémicamente en la planta, por lo que son llamados genes relacionados con la SAR. Otros genes que también gobiernan las reacciones de defensa no son expresados sistémicamente (Sha y Klessig, 1996).
.
Página siguiente: Inductores de defensa sintéticos
.
.
Bibliografía
Abramovitch RB, Anderson JC, Martin GB (2006) Bacterial elicitation and evasion of plant innate immunity. Nat. Rev. Mol. Cell Biol. 7: 601–611. doi: 10.1038/nrm1984
Agrios G (2005) Plant Pathology 5th Edition, eBook ISBN: 9780080473789
Ai G, Yang K, Ye W, et al. (2020) Prediction and characterization of RXLR effectors in Pythium species. Mol. Plant-Microbe Interact. 33: 1046-1058. doi: 10.1094/MPMI-01-20-0010-R
Andolfo G, Ercolano MR (2015) Plant Innate Immunity Multicomponent Model. Frontiers in Plant Science 6: 987. doi: 10.3389/fpls.2015.00987
Apoga D, Jansson H-B, Tunlid A (2001) Adhesion of conidia and germlings of the plant pathogenic fungus Bipolaris sorokiniana to solid surfaces. Mycological Research 105: 1251–1260. doi: 10.1016/S0953-7562(08)61997-8
Ayliffe MA, Lagudah ES (2004) Molecular genetics of disease resistance in cereals. Annals of Botany 94(6): 765-773. doi: 10.1093/aob/mch207
Balint-Kurti P (2019) The plant hypersensitive response: concepts, control and consequences. Molecular Plant Pathology 20: 1163-1178. doi: 10.1111/mpp.12821
Bari R, Jones JDG (2009) Role of plant hormones in plant defence responses. Plant Molecular Biology 69(4): 473–488. doi: 10.1007/s11103-008-9435-0
Barnes SN, Wram CL, Mitchum MG, Baum TJ (2018) The plant‐parasitic cyst nematode effector GLAND4 is a DNA‐binding protein. Molecular Plant Pathology. doi: 10.1111/mpp.12697
Bent AF, Mackey D (2007) Elicitors, Effectors, and R Genes: The New Paradigm and a Lifetime Supply of Questions. Annual Review of Phytopathology 45: 399-436. doi: 10.1146/annurev.phyto.45.062806.094427
Boller T (1995) Chemoperception of microbial signals in plant cells. Annual Review of Plant Physiology and Plant Molecular Biology 46: 189–214. doi: 10.1146/annurev.pp.46.060195.001201
Boller T, Felix G (2009) A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annual Review of Plant Biology 60: 379–406. doi: 10.1146/annurev.arplant.57.032905.105346
Bonardi V, Tang S, Stallmann A, et al. (2011) Expanded functions for a family of plant intracellular immune receptors beyond specific recognition of pathogen effectors. Proc. Natl. Acad. Sci. USA 108: 16 463–16 468. doi: 10.1073/pnas.1113726108
Bonas U, Lahaye T (2002) Plant disease resistance triggered by pathogen-derived molecules: refined models of specific recognition. Current Opinion in Microbiology 5: 44–50. doi: 10.1016/S1369-5274(02)00284-9
Burdett H, Bentham AR, Williams SJ, et al. (2019) The Plant “Resistosome”: Structural Insights into Immune Signaling. Cell Host & Microbe 26: 193-201. doi: 10.1016/j.chom.2019.07.020
Bernoux M, Zetzsche H, Stuttmann J (2022) Connecting the dots between cell surface- and intracellular-triggered immune pathways in plants. Current Opinion in Plant Biology 69: 102276. doi: 10.1016/j.pbi.2022.102276
Bjornson M, Pimprikar P, Nürnberger T, et al. (2021) The transcriptional landscape of Arabidopsis thaliana pattern-triggered immunity. Nat. Plants 7: 579–586. doi: 10.1038/s41477-021-00874-5
Carella P, Evangelisti E, Schornack S (2018) Sticking to it: phytopathogen effector molecules may converge on evolutionarily conserved host targets in green plants. Current Opinion in Plant Biology 44: 175-180. doi: 10.1016/j.pbi.2018.04.019
Cesari S (2018) Multiple strategies for pathogen perception by plant immune receptors. New Phytologist 219: 17-24. doi: 10.1111/nph.14877
Chiang YH, Coaker G (2015) Effector Triggered Immunity: NLR Immune Perception and Downstream Defense Responses. The Arabidopsis Book 13: e0183. doi: 10.1199/tab.0183
Chisholm ST, Coaker G, Day B, Staskawicz BJ (2006) Host-microbe interactions: shaping the evolution of the plant immune response. Cell 124(4): 803-814. doi: 10.1016/j.cell.2006.02.008
Christensen AB, Thordal-Christensen H, Zimmermann G, et al. (2004) The germinlike protein GLP4 exhibits superoxide dismutase activity and is an important component of quantitative resistance in wheat and barley. Molecular Plant-Microbe Interactions 17: 109–117. doi: 10.1094/MPMI.2004.17.1.109
Choudhary DK, Prakash A, Johri BN (2007) Induced systemic resistance (ISR) in plants: mechanism of action. Indian J Microbiol. 47(4): 289-297. doi: 10.1007/s12088-007-0054-2
Clark RM, Schweikert G, Toomajian C, et al. (2007) Common sequence polymorphisms shaping genetic diversity in Arabidopsis thaliana. Science 317: 338–342. doi: 10.1126/science.1138632
Cohn J, Sessa G, Martin GB (2001) Innate immunity in plants. Curr. Opin. Immunol. 13: 55–62. doi: 10.1016/S0952-7915(00)00182-5
Conrath U (2006) Systemic Acquired Resistance. Plant Signal Behav. 1(4):179-84. Link
D’Ovidio R, Mattei B, Roberti S, Bellincampi D (2004) Polygalacturonases, polygalacturonase-inhibiting proteins and pectic oligomers in plant–pathogen interactions. Biochimica et Biophysica Acta (BBA) – Proteins and Proteomics 1696: 237–244. doi: 10.1016/j.bbapap.2003.08.012
Dangl JL, Jones JD (2001) Plant pathogens and integrated defence responses to infection. Nature 411: 826-833. doi: 10.1038/35081161
Dangl JL, McDowell JM (2006) Two modes of pathogen recognition by plants. Proc. Natl. Acad. Sci. USA 103: 8575–8576. doi: 10.1073/pnas.0603183103
de Jonge R, Bolton MD, Thomma BP (2011) How filamentous pathogens co-opt plants: the ins and outs of fungal effectors. Current Opinion in Plant Biology 14(4): 400-6. doi: 10.1016/j.pbi.2011.03.005
De la Concepcion JC, Franceschetti M, Maqbool A, et al. (2018) Polymorphic residues in rice NLRs expand binding and response to effectors of the blast pathogen. Nature Plantsvolume 4: 576–585. doi: 10.1038/s41477-018-0194-x
De Lorenzo G, D’Ovidio R, Cervone F (2001) The role of polygalacturonase-inhibiting proteins (pgips) in defense against pathogenic fungi. Annual Review of Phytopathology 39: 313-335. doi: 10.1146/annurev.phyto.39.1.313
De Wit PJ, Mehrabi R, Van Den Burg HA, Stergiopoulos I (2009) Fungal effector proteins: past, present and future. Molecular Plant Pathology 10: 735-747. doi: 10.1111/j.1364-3703.2009.00591.x
De Wit PJGM (2016) Apoplastic fungal effectors in historic perspective; a personal view. New Phytol, 212: 805-813. doi: 10.1111/nph.14144
Dean JFD, Anderson JD (1991) Ethylene biosynthesis-inducing xylanase. II. Purification and physical characterization of the enzyme produced by Trichoderma viride. Plant Physiology 95: 316–323. doi: 10.1104/pp.95.1.316
Desjardins AE, Plattner RD (2000) Fumonisin B1-nonproducing strains of Fusarium verticillioides cause maize (Zea mays) ear infection and ear rot. Journal of Agricultural and Food Chemistry 48: 5773–5780. doi: 10.1021/jf000619k
Di Matteo A, Bonivento D, Tsernoglou D, et al. (2006) Polygalacturonase-inhibiting protein (PGIP) in plant defence: a structural view. Phytochemistry 67(6): 528-533. doi: 10.1016/j.phytochem.2005.12.025
Dickman MB, Ha Y-S, Yang B, et al. (2003) A protein kinase from Colletotrichum trifolii is induced by plant cutin and is required for appressorium formation. Molecular Plant-Microbe Interactions 16: 411–421. doi: 10.1094/MPMI.2003.16.5.411
Dickman MB, de Figueiredo P (2013) Death be not proud –cell death control in plant fungal interactions. PLoS Pathog. 9:e1003542. doi: 10.1371/journal.ppat.1003542
Dodds PN, Rathjen JP (2010) Plant immunity: towards an integrated view of plant–pathogen interactions. Nature Reviews Genetics 11: 539–548. doi: 10.1038/nrg2812
Dodds PN (2010) Plant science. Genome evolution in plant pathogens. Science 330(6010): 1486-7. doi: 10.1126/science.1200245
Dreischhoff S, Das IS, Jakobi M, et al. (2020) Local Responses and Systemic Induced Resistance Mediated by Ectomycorrhizal Fungi. Front. Plant Sci. 11: 590063. doi: 10.3389/fpls.2020.590063
Duggan C, Moratto E, Savage Z, et al. (2021) Dynamic localization of a helper NLR at the plant–pathogen interface underpins pathogen recognition. Proceedings of the National Academy of Sciences 118 (34): e2104997118. doi: 10.1073/pnas.2104997118
Duxbury Z, Ma Y, Furzer OJ, et al. (2016) Pathogen perception by NLRs in plants and animals: parallel worlds. BioEssays 38: 769–781. doi: 10.1002/bies.201600046
Eastman S, Bayless A, Guo M (2022) The Nucleotide Revolution: Immunity at the Intersection of Toll/Interleukin-1 Receptor Domains, Nucleotides, and Ca2. Mol Plant Microbe Interact. 35(11): 964-976. doi: 10.1094/MPMI-06-22-0132-CR
El Kasmi F (2021) How activated NLRs induce anti-microbial defenses in plants. Biochem Soc Trans BST20210242. doi: 10.1042/BST20210242
Ellis JG, Lawrence GJ, Luck JE, Dodds PN (1999) Identification of regions in alleles of the flax rust resistance gene L that determine differences in gene-for-gene specificity. Plant Cell 11: 495–506. doi: 10.1105/tpc.11.3.495
(2011) Showdown at the RXLR motif: Serious differences of opinion in how effector proteins from filamentous eukaryotic pathogens enter plant cells. Proc. Natl. Acad. Sci. U.S.A. 108: 14381-14382. doi: 10.1073/pnas.1111668108
Fabro G (2021) Oomycete intracellular effectors: specialized weapons targeting strategic plant processes. New Phytol. doi: 10.1111/nph.17828
Fernández-Milmanda GL (2023) Rise of signaling: jasmonic and salicylic acid oppositely control reactive oxygen species wave production. Plant Physiol. 191(2): 814-816. doi: 10.1093/plphys/kiac517
Fluhr R (2001) Sentinels of disease. Plant resistance genes. Plant Physiologist 127: 1367–1374. doi: 10.1104/pp.010763
Fritz-Laylin LK, Krishnamurthy N, Tor M, et al. (2005) Phylogenomic analysis of the receptor-like proteins of rice and Arabidopsis. Plant Physiol 138: 611–623. doi: 10.1104/pp.104.054452
Fujita K, Wright AJ, Meguro A, et al. (2004) Rapid pregermination and germination responses of Erysiphe pisi conidia to contact and light. Journal of General Plant Pathology 70: 75–84. doi: 10.1007/s10327-003-0093-4
(2014) Bacterial type III secretion systems: Specialized nanomachines for protein delivery into target cells. Annu. Rev. Microbiol. 68: 415-438. doi: 10.1146/annurev-micro-092412-155725
Gao M, He Y, Yin X, et al. (2021) Ca2+ sensor-mediated ROS scavenging suppresses rice immunity and is exploited by a fungal effector. Cell 184: 5391-5404.e17. doi: 10.1016/j.cell.2021.09.009
Garnica DP, Nemri A, Upadhyaya NM, et al. (2014) The Ins and Outs of Rust Haustoria. PLoS Pathog 10(9): e1004329. doi: 10.1371/journal.ppat.1004329
Gasperini D, Howe GA (2024) Phytohormones in a universe of regulatory metabolites: Lessons from jasmonate. Plant Physiol.: kiae045. doi: 10.1093/plphys/kiae045
Gassmann W, Bhattacharjee S (2012) Effector-Triggered Immunity Signaling: From Gene-for-Gene Pathways to Protein-Protein Interaction Networks. Molecular Plant-Microbe Interactions 25(7): 862-868. doi: 10.1094/MPMI-01-12-0024-IA
Giraldo MC, Valent B (2013) Filamentous plant pathogen effectors in action. Nature Reviews Microbiology 11: 800–814. doi: 10.1038/nrmicro3119
(2013) Two distinct secretion systems facilitate tissue invasion by the rice blast fungus Magnaporthe oryzae. Nat. Commun. 4: 1996. doi: 10.1038/ncomms2996
Gish LA, Clark SE (2011) The RLK/Pelle family of kinases. The Plant Journal 66: 117-127. doi: 10.1111/j.1365-313X.2011.04518.x
Gjetting T, Carver TLW, Skøt L, Lyngær MF (2004) Differential gene expression in individual papilla-resistant and powdery mildew-infected barley epidermal cells. Molecular Plant–Microbe Interactions 17: 729–738. doi: 10.1094/MPMI.2004.17.7.729
Glazebrook J, Chen W, Estes B, et al. (2003) Topology of the network integrating salicylate and jasmonate signal transduction derived from global expression phenotyping. Plant J. 34: 217–228. doi: 10.1046/j.1365-313X.2003.01717.x
Goellner K, Conrath U (2008) Priming: it’ s all the world to induced disease resistance. European Journal of Plant Pathology 121(3): 233–242. doi 10.1007/s10658-007-9251-4
Grant M, Lamb C (2006) Systemic immunity. Current Opinion in Plant Biology 9(4): 414–420. doi: 10.1016/j.pbi.2006.05.013
Haas BJ, Kamoun S, Zody MC, et al. (2009) Genome sequence and analysis of the Irish potato famine pathogen Phytophthora infestans. Nature 461: 393–398. doi: 10.1038/nature08358
Hammerschmidt R (1999) Induced disease resistance: how do induced plants stop pathogens? Physiological and Molecular Plant Pathology 55(2): 77-84. doi: 10.1006/pmpp.1999.0215
Hammerschmidt R (2004) Myriad molecular resistance signals: conidial, endogenous and inorganic. Physiological and Molecular Plant Pathology 64: 165–167. doi: 10.1016/j.pmpp.2004.11.003
He Y, Zhou J, Shan L, Meng X (2018) Plant cell surface receptor-mediated signaling – a common theme amid diversity. J Cell Sci 131: jcs209353. doi: 10.1242/jcs.209353
Heath MC (2000) Nonhost resistance and nonspecific plant defenses. Current Opinion in Plant Biology 3(4): 315-319. doi: 10.1016/S1369-5266(00)00087-X
Henry E, Toruño TY, Jauneau A, et al. (2017) Direct and Indirect Visualization of Bacterial Effector Delivery into Diverse Plant Cell Types during Infection. The Plant Cell 29(7): 1555-1570. doi: 10.1105/tpc.17.00027
Jacob F, Vernaldi S, Maekawa T (2013) Evolution and conservation of plant NLR functions. Frontiers in Immunology 4: 297. doi: 10.3389/fimmu.2013.00297
Jamieson PA, Shan L, He P (2018) Plant cell surface molecular cypher: Receptor-like proteins and their roles in immunity and development. Plant Science 274: 242-251. doi: 10.1016/j.plantsci.2018.05.030
Jia A, Huang S, Ma S, et al. (2023) TIR-catalyzed nucleotide signaling molecules in plant defense. Current Opinion in Plant Biology 73: 102334. doi: 10.1016/j.pbi.2022.102334
Jin H, Choi SM, Kang MJ, et al. (2018) Salicylic acid-induced transcriptional reprogramming by the HAC–NPR1–TGA histone acetyltransferase complex in Arabidopsis. Nucleic Acids Research gky847. doi: 10.1093/nar/gky847
Jones D, Thomas C, Hammond-Kosack K, et al. (1994) Isolation of the tomato Cf-9 gene for resistance to Cladosporium fulvum by transposon tagging. Science 266: 789–793. doi: 10.1126/science.7973631
Jones JDG, Dangl JL (2006) The plant immune system. Nature 444: 323–329. doi: 10.1038/nature05286
Jones JD, Vance RE, Dangl JL (2016) Intracellular innate immune surveillance devices in plants and animals. Science, 354, aaf6395. doi: 10.1126/science.aaf6395
Jung HW, Tschaplinski TJ, Wang L, et al. (2009) Priming in systemic plant immunity. Science 324: 89–91. doi: 10.1126/science.1170025
Kabbage M, Yarden O, Dickman MB (2015) Pathogenic attributes of Sclerotinia sclerotiorum: switching from a biotrophic to necrotrophic lifestyle. Plant Sci. 233: 53–60. doi: 10.1016/j.plantsci.2014.12.018
Kagan J (2023) Infection infidelities drive innate immunity. Science 379, Issue 6630: 333-335. doi: 10.1126/science.ade9733
Kato H, Nemoto K, Shimizu M, et al. (2021) Pathogen-derived 9-methyl sphingoid base is perceived by a lectin receptor kinase in Arabidopsis. bioRxiv 2021.10.18.464766; doi: 10.1101/2021.10.18.464766
Keen NT, Partridge JE, Zaki AI (1972) Pathogen-produced elicitor of a chemical defense mechanism in soybeans monogenically resistant to Phytophthora megasperma var. sojae. Phytopathology 62: 768.
(2010) Translocation of Magnaporthe oryzae effectors into rice cells and their subsequent cell-to-cell movement. Plant Cell 22: 1388-1403. doi: 10.1105/tpc.109.069666
Khokhani D, Carrera Carriel C, Vayla S, et al. (2021) Deciphering the Chitin Code in Plant Symbiosis, Defense, and Microbial Networks. Annu Rev Microbiol. 75: 583-607. doi: 10.1146/annurev-micro-051921-114809
Klessig DF, Choi HW, Dempsey DA (2018) Systemic Acquired Resistance and Salicylic Acid: Past, Present, and Future. Molecular Plant-Microbe Interactions. doi: 10.1094/MPMI-03-18-0067-CR
Klusener B, Weiler EW (1999) Pore-forming properties of elicitors of plant defense reactions and cellulolytic enzymes. FEBS Lett. 459: 263–266. doi: 10.1016/S0014-5793(99)01261-2
Kourelis J, Marchal C, Kamoun S (2021) NLR immune receptor-nanobody fusions confer plant disease resistance. bioRxiv 2021.10.24.465418; doi: 10.1101/2021.10.24.465418
Krasileva KV, Dahlbeck D, Staskawicz BJ (2010) Activation of an Arabidopsis resistance protein is specified by the in planta association of its leucine-rich repeat domain with the cognate oomycete effector. Plant Cell 22: 2444–58. doi: 10.1105/tpc.110.075358
Kruger WM, Szabo LJ, Zeyen RJ (2003) Transcription of the defense response genes chitinase IIb, PAL, and peroxidase is induced by the barley powdery mildew fungus and is only indirectly modulated by R genes. Physiological and Molecular Plant Pathology 63: 167–178. doi: 10.1016/j.pmpp.2003.10.006
Kruijt M, De Kock MJ, De Wit PJ (2005) Receptor‐like proteins involved in plant disease resistance. Molecular Plant Pathology 6: 85-97. doi: 10.1111/j.1364-3703.2004.00264.x
Lai Z, Wang F, Zheng Z, et al. (2011) A critical role of autophagy in plant resistance to necrotrophic fungal pathogens. The Plant Journal 66: 953–968. doi: 10.1111/j.1365-313X.2011.04553.x
Lai, Y, Eulgem T (2018) Transcript‐level expression control of plant NLR genes. Molecular Plant Pathology 19: 1267-1281. doi: 10.1111/mpp.12607
Lee HA, Lee HY, Seo E, et al. (2017) Current Understandings of Plant Nonhost Resistance. Mol Plant Microbe Interact. 30(1): 5-15. doi: 10.1094/MPMI-10-16-0213-CR
Leary AY, Sanguankiattichai N, Duggan C, et al. (2018) Modulation of plant autophagy during pathogen attack. Journal of Experimental Botany 69(6): 1325–1333. doi: 10.1093/jxb/erx425
Lehti-Shiu MD, Zou C, Hanada K, Shiu SH (2009) Evolutionary history and stress regulation of plant receptor-like kinase/pelle genes. Plant Physiol. 150: 12–26. doi: 10.1104/pp.108.134353
Lev S, Horwitz BA (2003) A mitogen-activated protein kinase pathway modulates the expression of two cellulase genes in Cochliobolus heterostophus during plant infection. The Plant Cell 15: 835–844. doi: 10.1105/tpc.010546
Li L, Xue C, Bruno K, et al. (2004) Two PAK kinase genes, CHM1 and MST20, have distinct functions in Magnaporthe grisea. Molecular Plant-Microbe Interactions 17: 547–556. doi: 10.1094/MPMI.2004.17.5.547
Li X, Kapos P, Zhang Y (2015) NLRs in plants. Current Opinion in Immunology 32: 114–121. doi: 10.1016/j.coi.2015.01.014
Li B, Meng X, Shan L, He P (2016) Transcriptional Regulation of Pattern-Triggered Immunity in Plants. Cell Host Microbe. 19(5): 641-50. doi: 10.1016/j.chom.2016.04.011
Li F, Wang A (2018) RNA decay is an antiviral defense in plants that is counteracted by viral RNA silencing suppressors. PLoS Pathog 14(8): e1007228. doi: 10.1371/journal.ppat.1007228
Li L, Habring A, Wang K, Weigel D (2020) Atypical resistance protein RPW8/HR triggers oligomerization of the NLR immune receptor RPP7 and autoimmunity. Cell Host Microbe 27, 405–417.e6. doi: 10.1016/j.chom.2020.01.012
Liu PL, Du L, Huang Y, et al. (2017) Origin and diversification of leucine-rich repeat receptor-like protein kinase (LRR-RLK) genes in plants. BMC Evol Biol. 17: 47. doi: 10.1186/s12862-017-0891-5
Liu Y, Zhang X, Yuan G, et al. (2021) A designer rice NLR immune receptor confers resistance to the rice blast fungus carrying noncorresponding avirulence effectors. PNAS 118 (44): e2110751118. doi: 10.1073/pnas.2110751118
(2017) How filamentous plant pathogen effectors are translocated to host cells. Curr. Opin. Plant Biol. 38: 19-24. doi: 10.1016/j.pbi.2017.04.005
Lu H, Higgins VJ (1998) Measurement of active oxygen species generated in planta in response to elicitor AVR9 of Cladosporium fulvum. Physiological and Molecular Plant Pathology 52: 35–51. doi: 10.1006/pmpp.1997.0138
Luderer R, Rivas S, Nurnberger T, et al. (2001) No evidence for binding between resistance gene product Cf-9 of tomato and avirulence gene product AVR9 of Cladosporium fulvum. Mol. Plant. Microbe Interact. 14 867–876. doi: 10.1094/MPMI.2001.14.7.867
Luderer R, Joosten MHAJ (2001) Avirulence proteins of plant pathogens: determinants of victory and defeat. Molecular Plant Pathology 2: 355–364. doi: 10.1046/j.1464-6722.2001.00086.x
(2021) A cell surface-exposed protein complex with an essential virulence function in Ustilago maydis. Nat. Microbiol. 6: 722-730. doi: 10.1038/s41564-021-00896-x
Ma S, Lapin D, Liu L, et al. (2020) Direct pathogen-induced assembly of an NLR immune receptor complex to form a holoenzyme. Science 370: eabe3069. doi: 10.1126/science.abe3069
Maekawa T, Kufer T, Schulze-Lefert P (2011) NLR functions in plant and animal immune systems: so far and yet so close. Nat Immunol 12: 817–826. doi: 10.1038/ni.2083
Mandadi KK, Scholthof KB (2013) Plant immune responses against viruses: how does a virus cause disease? Plant Cell 25(5): 1489-505. doi: 10.1105/tpc.113.111658
Manning VA, Ciuffetti LM (2005) Localization of Ptr ToxA produced by Pyrenophora tritici-repentis reveals protein import into wheat mesophyll cells. The Plant Cell 17: 3203–3212. doi: 10.1105/tpc.105.035063
Marchal C, Zhang J, Zhang P, et al. (2018) BED-domain-containing immune receptors confer diverse resistance spectra to yellow rust. Nature Plants. doi: 10.1038/s41477-018-0236-4
Martin R, Qi T, Zhang H, et al. (2020) Structure of the activated ROQ1 resistosome directly recognizing the pathogen effector xopQ. Science 370: eabd9993. doi: 10.1126/science.abd9993
Mayer MG, Ziegler E (1988) An elicitor from Phytophthora megasperma f. sp. glycinea influences the membrane potential of soybean cotyledonary cells. Physiological and Molecular Plant Pathology 33(3): 397-407. doi: 10.1016/0885-5765(88)90006-9
Mbengue M, Navaud O, Peyraud R, et al. (2016) Emerging Trends in Molecular Interactions between Plants and the Broad Host Range Fungal Pathogens Botrytis cinerea and Sclerotinia sclerotiorum. Frontiers in Plant Science 7: 422. doi: 10.3389/fpls.2016.00422
Meyers BC, Kozik A, Griego A, et al. (2003) Genome wide analysis of NBS-LRR-encoding genes in Arabidopsis. Plant Cell 15: 809–834. doi: 10.1105/tpc.009308
(2013) Nematode effector proteins: An emerging paradigm of parasitism. New Phytol. 199: 879-894. doi: 10.1111/nph.12323
Monaghan J, Zipfel C (2012) Plant pattern recognition receptor complexes at the plasma membrane. Current Opinion in Plant Biology 15(4): 349-357. doi: 10.1016/j.pbi.2012.05.006
Monjil MS, Kato H, Matsuda K, et al. (2021) Two structurally different oomycete MAMPs induce distinctive plant immune responses. bioRxiv 2021.10.22.465218. doi: 10.1101/2021.10.22.465218
Monteiro F, Nishimura MT (2018) Structural, Functional, and Genomic Diversity of Plant NLR Proteins: An Evolved Resource for Rational Engineering of Plant Immunity. Annual Review of Phytopathology 56: 12.1–12.25. doi: 10.1146/annurev-phyto-080417-045817
Montesano M, Brader G, Palva ET (2003) Pathogen-derived elicitors: searching for receptors in plants. Molecular Plant Pathology 4: 73–78. doi: 10.1046/j.1364-3703.2003.00150.x
Mould MJR, Xu T, Barbara M, et al. (2003) cDNAs generated from individual epidermal cells reveal that differential gene expression predicting subsequent resistance or susceptibility to rust fungal infection occurs prior to the fungus entering the cell lumen. Molecular Plant-Microbe Interactions 16: 835–845. doi: 10.1094/MPMI.2003.16.9.835
Nandi A, Kachroo P, Fukushige H, et al. (2003) Ethylene and jasmonic acid signaling affect the NPR1-independent expression of defense genes without impacting resistance to Pseudomonas syringae and Peronospora parasitica in the Arabidopsis ssi1 mutant. Molecular Plant-Microbe Interactions 18: 588–599. doi: 10.1094/MPMI.2003.16.7.588
Neu C, Keller B, Feuillet C (2003) Cytological and molecular analysis of the Hordeum vulgare–Puccinia triticina nonhost interaction. Molecular Plant-Microbe Interactions 16: 626–633. doi: 10.1094/MPMI.2003.16.7.626
Ngou BPM, Ahn HK, Ding P, et al. (2021) Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature 592: 110–115. doi: 10.1038/s41586-021-03315-7
Nielsen KA, Nicholson RL, Carver TLW, et al. (2000) First touch: an immediate response to signal recognition in conidia of Blumeria graminis. Physiological and Molecular Plant Pathology 56: 63–70. doi: 10.1006/pmpp.1999.0241
Nimchuk Z, Rohmer L, Chang JH, Dangl JL (2001) Knowing the dancer from the dance: R-gene products and their interactions with other proteins from host and pathogen. Current Opinion in Plant Biology 4: 288–294. doi: 10.1016/S1369-5266(00)00175-8
Nishad R, Ahmed T, Rahman VJ, Kareem A (2020) Modulation of Plant Defense System in Response to Microbial Interactions. Front. Microbiol. 11: 1298. doi: 10.3389/fmicb.2020.01298
Nurnberger T, Scheel D (2001) Signal transmission in the plant immune response. Trends in Plant Science 6: 372–379. doi: 10.1016/S1360-1385(01)02019-2
Ossowski S, Schneeberger K, Clark RM, et al. (2008) Sequencing of natural strains of Arabidopsis thaliana with short reads. Genome Res. 18: 2024–2033. doi: 10.1101/gr.080200.108
Panstruga R, Moscou MJ (2020) What is the Molecular Basis of Nonhost Resistance? Mol Plant Microbe Interact. 33(11): 1253-1264. doi: 10.1094/MPMI-06-20-0161-CR
Pi L, Yin Z, Duan W, et al. (2021) A G-type lectin receptor-like kinase regulates the perception of oomycete apoplastic expansin-like proteins. J Integr Plant Biol. doi: 10.1111/jipb.13194
Pieterse CMJ, Leon-Reyes A, Van der Ent S, Van Wees SCM (2009) Networking by small-molecule hormones in plant immunity. Nature Chemical Biology 5: 308–316. doi: 10.1038/nchembio.164
Pritchard L, Birch PR (2014) Zigzag model of plant–microbe interactions. Molecular Plant Pathology 15: 865-870. doi: 10.1111/mpp.12210
Richard MM, Gratias A, Meyers BC, Geffroy V (2018) Molecular mechanisms that limit the costs of NLR‐mediated resistance in plants. Molecular Plant Pathology. doi:10.1111/mpp.12723
Rivas S, Genin S (2011) A plethora of virulence strategies hidden behind nuclear targeting of microbial effectors. Frontiers in Plant Science 2: 104. doi: 10.3389/fpls.2011.00104
(2013) Toward understanding the role of aphid effectors in plant infestation. Mol. Plant-Microbe Interact. 26: 25-30. doi: 10.1094/MPMI-05-12-0119-FI
Ryerson DE, Heath MC (1992) Fungal elicitation of wall modifications in leaves of Phaseolus vulgaris L. cv. Pinto II. Effects of fungal wall components. Physiological and Molecular Plant Pathology 40: 283–298. doi: 10.1016/0885-5765(92)90077-9
Sacristán S, Goss EM, Eves-van den Akker S (2021) How Do Pathogens Evolve Novel Virulence Activities? Mol Plant Microbe Interact. MPMI09200258IA. doi: 10.1094/MPMI-09-20-0258-IA
Sánchez-Vallet A, Fouché S, Fudal I, et al. (2018) The Genome Biology of Effector Gene Evolution in Filamentous Plant Pathogens. Annual Review of Phytopathology 56. doi: 10.1146/annurev-phyto-080516-035303
Schneider M, Schweizer P, Meuwly P, Métraux JP (1996) Systemic Acquired Resistance in Plants. International Review of Cytology 168: 303-340. doi: 10.1016/S0074-7696(08)60887-6
(2010) Ancient class of translocated oomycete effectors targets the host nucleus. Proc. Natl. Acad. Sci. U.S.A. 107: 17421-17426. doi: 10.1073/pnas.1008491107
Selin C, de Kievit TR, Belmonte MF, Fernando WGD (2016) Elucidating the Role of Effectors in Plant-Fungal Interactions: Progress and Challenges. Front. Microbiol. 7: 600. doi: 10.3389/fmicb.2016.00600
Shah J, Klessig DF (1996) Identification of a salicylic acid‐responsive element in the promoter of the tobacco pathogenesis‐related β‐1,3‐glucanase gene, PR‐2d. The Plant Journal 10: 1089-1101. doi: 10.1046/j.1365-313X.1996.10061089.x
Shiu SH, Bleecker AB (2001a) Plant receptor-like kinase gene family: diversity, function, and signaling. Science’s STKE 18(113): re22. doi: 10.1126/stke.2001.113.re22
Shiu SH, Bleecker AB (2001b) Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. U. S.A. 98: 10763–10768. doi: 10.1073/pnas.181141598
Shiu SH, Bleecker AB (2003) Expansion of the receptor-like kinase/pelle gene family and receptor-like proteins in Arabidopsis. Plant Physiol. 132: 530–543. doi: 10.1104/pp.103.021964
Shlezinger N, Minz A, Gur Y, et al. (2011) Anti-apoptotic machinery protects the necrotrophic fungus Botrytis cinerea from host-induced apoptotic-like cell death during plant infection. PLoS Pathog. 7:e1002185. doi: 10.1371/journal.ppat.1002185
Sperschneider J, Dodds PN (2022) EffectorP 3.0: Prediction of Apoplastic and Cytoplasmic Effectors in Fungi and Oomycetes. Mol Plant Microbe Interact. 35(2): 146-156. doi: 10.1094/MPMI-08-21-0201-R
Steele JFC, Hughes RK, Banfield MJ (2019) Structural and biochemical studies of an NB-ARC domain from a plant NLR immune receptor. PLoS ONE 14(8): e0221226. doi: 10.1371/journal.pone.0221226
Stergiopoulos I, de Wit PJ (2009) Fungal effector proteins. Annual Review of Phytopathology 47: 233-263. doi: 10.1146/annurev.phyto.112408.132637
Stotz HU, Mitrousia GK, de Wit P, Fitt BDL (2014) Effector-triggered defence against apoplastic fungal pathogens. Trends Plant Sci. 19: 491–500. doi: 10.1016/j.tplants.2014.04.009
(2021) Cell wall-associated effectors of plant-colonizing fungi. Mycologia 113: 247-260. doi: 10.1080/00275514.2020.1831293
Thaler JS, Owen B, Higgins VJ (2004) The role of the jasmonate response in plant susceptibility to diverse pathogens with a range of lifestyles. Plant Physiology 135: 530–538. doi: 10.1104/pp.104.041566
Thieffry A, López-Márquez D, Bornholdt J, et al. (2022) PAMP-triggered genetic reprogramming involves widespread alternative transcription initiation and an immediate transcription factor wave. The Plant Cell 34: 2615–2637. doi: 10.1093/plcell/koac108
Thilmony R, Underwood W, He SY (2006) Genome-wide transcriptional analysis of the Arabidopsis thaliana interaction with the plant pathogen Pseudomonas syringae pv. tomato DC3000 and the human pathogen Escherichia coli O157:H7. Plant J. 46(1): 34-53. Erratum in: Plant J. 2019 98(6): 1158. doi: 10.1111/j.1365-313X.2006.02725.x
Thomas CM, Jones DA, Parniske M, et al. (1997) Characterization of the tomato Cf-4 gene for resistance to Cladosporium fulvum identifies sequences that determine recognitional specificity in Cf-4 and Cf-9. Plant Cell 9: 2209–2224. doi: 10.1105/tpc.9.12.2209
Tian D, Traw MB, Chen JQ, et al. (2003) Fitness costs of R-gene-mediated resistance in Arabidopsis thaliana. Nature 423: 74–77. doi: 10.1038/nature01588
Tian H, Chen S, Wu Z, et al (2021) Activation of TIR signaling is required for pattern-triggered immunity. bioRxiv. doi: 10.1101/2020.12.27.424494
Tör M, Lotze MT, Holton N (2009) Receptor-mediated signalling in plants: molecular patterns and programmes. Journal of Experimental Botany 60(13): 3645–3654. doi: 10.1093/jxb/erp233
Tsuda K, Somssich IE (2015) Transcriptional networks in plant immunity. New Phytol 206: 932-947. doi: 10.1111/nph.13286
Tsuji G, Fujii S, Tsuge S, et al. (2003) The Colletotrichum lagenarium Stel 2-like gene CST1 is essential for appressorium penetration. Mol. Plant Microbe Interact. 167: 315–325.
Tyler BM, Tripathy S, Zhang X, et al. (2006) Phytophthora genome sequences uncover evolutionary origins and mechanisms of pathogenesis. Science 313: 1261–1266. doi: 10.1126/science.1128796
Uppalapati SR, Toyoda K, Yasuhiro I, et al. (2004) Differential regulation of MBP kinases by a glycoprotein elicitor and a polypeptide suppressor from Mycosphaerella pinodes in pea. Physiological and Molecular Plant Pathology 64: 17–25. doi: 10.1016/j.pmpp.2004.05.003
Vallad GE, Goodman RM (2004) Systemic Acquired Resistance and Induced Systemic Resistance in Conventional Agriculture. Crop Science 44: 1920-1934. doi: 10.2135/cropsci2004.1920
Van der Biezen EA, Jones JD (1998) Plant disease-resistance proteins and the gene-for-gene concept. Trends in Biochemical Sciences 23: 454–456. doi: 10.1016/S0968-0004(98)01311-5
Vidhyasekaran P (1988) Physiology of Disease Resistance in Plants. Vol. I, CRC Press, Boca Raton, FL, p. 149.
Vidhyasekaran P (2007) Fungal Pathogenesis in Plants and Crops. Molecular Biology and Host Defense Mechanisms. CRC Press, Second Edition, 536 p. eBook ISBN 9781420021035
Voegele RT, Mendgen K (2003) Rust haustoria: nutrient uptake and beyond. New Phytologist 159: 93-100. doi: 10.1046/j.1469-8137.2003.00761.x
Wan L, He Z (2021) NADase and now Ca2+ channel, what else to learn about plant NLRs?. Stress Biology 1: 7. doi: 10.1007/s44154-021-00007-0
Wang G, Ellendorff U, Kemp B, et al. (2008) A genome-wide functional investigation into the roles of receptor-like proteins in Arabidopsis. Plant Physiol. 147: 503–517. doi: 10.1104/pp.108.119487
Wang Q, Han C, Ferreira AO, et al. (2011) Transcriptional programming and functional interactions within the Phytophthora sojae RXLR effector repertoire. Plant Cell. 23(6): 2064-2086. doi: 10.1105/tpc.111.086082
Wang J, Hu M, Wang J, et al. (2019a) Reconstitution and structure of a plant NLR resistosome conferring immunity. Science 364: eaav5870. doi: 10.1126/science.aav5870
Wang J, Wang J, Hu M, et al. (2019b) Ligand-triggered allosteric ADP release primes a plant NLR complex. Science 364:eaav5868. doi: 10.1126/science.aav5868
Wang W, Chen L, Fengler K, et al. (2021) A giant NLR gene confers broad-spectrum resistance to Phytophthora sojae in soybean. Nat Commun. 12(1): 6263. doi: 10.1038/s41467-021-26554-8
Wang S, Boevink PC, Welsh L, et al. (2017) Delivery of cytoplasmic and apoplastic effectors from Phytophthora infestans haustoria by distinct secretion pathways. New Phytol, 216: 205-215. doi: 10.1111/nph.14696
(2017) The RxLR motif of the host targeting effector AVR3a of Phytophthora infestans is cleaved before secretion. Plant Cell 29: 1184-1195. doi: 10.1105/tpc.16.00552
(2007) A translocation signal for delivery of oomycete effector proteins into host plant cells. Nature 450: 115-118. doi: 10.1038/nature06203
(2020) Effector prediction and characterization in the oomycete pathogen Bremia lactucae reveal host-recognized WY domain proteins that lack the canonical RXLR motif. PLoS Pathog. 16: e1009012. doi: 10.1371/journal.ppat.1009012
Wright AJ, Carver TLW, Thomas BJ, et al. (2000) The rapid and accurate determination of germ tube emergence site by Blumeria graminis conidia. Physiological and Molecular Plant Pathology 57: 281–301. doi: 10.1006/pmpp.2000.0304
Wu C-H, Abd-El-Haliem A, Bozkurt TO, et al. (2017) NLR network mediates immunity todiverse plant pathogens. Proceedings of the National Academy ofSciences, USA 114: 8113–8118. doi: 10.1073/pnas.1702041114
Yamaoka N, Lyons PC, Hipskind J, Nicholson RL (1990) Elicitor of sorghum phytoalexin synthesis from Colletotrichum graminicola. Physiological and Molecular Plant Pathology 37: 255–270. doi: 10.1016/0885-5765(90)90075-9
Yang Q, Trinh HX, Imai S, et al. (2004) Analysis of the involvement of hydroxyanthranilate hydroxycinnamoyltransferase and caffeoyl-CoA 3-O-methyltransferase in phytoalexin biosynthesis in oat. Molecular Plant-Microbe Interactions 17: 81–89. doi: 10.1094/MPMI.2004.17.1.81
Yang Z, Rogers LM, Song Y, et al. (2005) Homoserine and asparagine are host signals that trigger in planta expression of a pathogenesis gene in Nectria haematococca. PNAS 102: 4197–4202. doi: 10.1073/pnas.0500312102
Yu TY, Sun MK, Liang LK (2021) Receptors in the Induction of the Plant Innate Immunity. Mol Plant Microbe Interact. MPMI07200173CR. doi: 10.1094/MPMI-07-20-0173-CR
, (2021) Long-distance communication through systemic macromolecular signaling mediates stress defense responses in plants. Physiologia Plantarum, 1– 9. doi: 10.1111/ppl.13535
Zhang Y, Tang M, Huang M, et al. (2022) Dynamic enhancer transcription associates with reprogramming of immune genes during pattern triggered immunity in Arabidopsis. BMC Biol 20: 165. doi: 10.1186/s12915-022-01362-8
Zhong M, Yan H, Li Y (2017) Flagellin: a unique microbe-associated molecular pattern and a multi-faceted immunomodulator. Cell Mol Immunol 14: 862–864. doi: 10.1038/cmi.2017.78
Zipfel C (2008) Pattern-recognition receptors in plant innate immunity. Current Opinion in Immunology 20(1): 10-16. doi: 10.1016/j.coi.2007.11.003
Zipfel C (2014) Plant pattern-recognition receptors. Trends in Immunology 35(7): 345-351. doi: 10.1016/j.it.2014.05.004